Vakgroep Biologie
Onderzoeksgroep Terrestrial Ecology Unit (Terec)
_____________________________________________________________________________________
DE WEZEL (MUSTELA NIVALIS) EN DE
HERMELIJN (MUSTELA ERMINEA) IN
VLAANDEREN
KENNISVERGARING AAN DE AUTOPSIETAFEL
Timo Van der Veken
Studentennummer: 01005009Promotor(en): prof. dr. Maurice Hoffmann
Wetenschappelijk begeleider:
Koen Van Den Berge
Masterproef voorgelegd voor het behalen van de graad van Master of Science in Biology Academiejaar: 2019 - 2020
© Faculty of Sciences – Onderzoeksgroep Terrestrial Ecology Unit (Terec)
All rights reserved. This thesis contains confidential information and confidential research results that are property to the UGent. The contents of this master thesis may under no circumstances be made public, nor complete or partial, without the explicit and preceding permission of the UGent representative, i.e. the supervisor. The thesis may under no circumstances be copied or duplicated in any form, unless permission granted in written form. Any violation of the confidential nature of this thesis may impose irreparable damage to the UGent. In case of a dispute that may arise within the context of this declaration, the Judicial Court of Gent only is competent to be notified.
Deze masterproef bevat vertrouwelijke informatie en vertrouwelijke onderzoeksresultaten die toebehoren aan de UGent. De inhoud van de masterproef mag onder geen enkele manier publiek gemaakt worden, noch geheel noch gedeeltelijk zonder de uitdrukkelijke schriftelijke voorafgaandelijke toestemming van de UGent-vertegenwoordiger, in casu de promotor. Zo is het nemen van kopieën of het op eender welke wijze dupliceren van het eindwerk verboden, tenzij met schriftelijke toestemming. Het niet respecteren van de confidentiële aard van het eindwerk veroorzaakt onherstelbare schade aan de UGent. Ingeval een geschil zou ontstaan in het kader van deze verklaring, zijn de rechtbanken van het arrondissement Gent uitsluitend bevoegd daarvan kennis te nemen.
1
Dankwoord
Het schrijven van mijn masterproef is een leerrijk proces geweest. De laatste 2 jaar heeft mij enorm veel bijgebracht omtrent de biologische onderzoekswereld.
Het afwerken van dit onderzoek, zou mij niet gelukt zijn zonder de hulp van een aantal belangrijke mensen. In eerste instantie, wil ik graag prof. Dr. Hofmann M. bedanken voor het mogelijk maken van dit thesisonderwerp, als promotor en mij deze unieke ervaring aan te bieden.
Vervolgens, wil ik graag mijn begeleider, Koen Van Den Berge, bedanken voor het mogelijk maken van dit thesisonderwerp en het beantwoorden van mijn vragen. Niet alleen omtrent dit thesisonderwerp, maar rond al mijn vragen van het carnivorenonderzoek. Daarnaast wil ik ook mijn dankbaarheid uitdrukken om een bijdrage te mogen leveren aan het wolvenproject als haaronderzoeker. Dit is een ervaring die ik nooit zal vergeten.
Voorts wil ik ook Jan Gouwy bedanken, omdat ik ook bij hem terechtkon voor al mijn vragen. De vondst van de “wolvenkill” in Leopoldsburg is een van de hoogtepunten geweest van de afgelopen 2 jaar. Ook wil ik mijn dank betuigen aan Filip Berlengee om mij de knepen van het vak te leren en om steeds een steun te zijn in moeilijke tijden. Ik bewonder zijn engagement, zowel op de werkvloer als in het VOC. Verder, wil ik ook zeker de andere werknemers in Geraardsbergen, met Kurt Schamp in het bijzonder, niet vergeten en wil ik hen bedanken voor de fijne tijd, die we samen hebben doorgebracht.
Ook Diemer Vercavie en Kristijn Swinnen, van de dienst Studie van Natuurpunt, zijn bedankt voor hun begeleiding.
Voorts, wil ik mijn beste vrienden, Floris, Ruben, Glen en Virginie bedanken voor de vriendschap en om er steeds voor mij te zijn.
Tot slot, wil ik mijn ouders een hart onder de riem stoppen. Het was geen evidente keuze om na mijn bachelor psychologie, over te schakelen naar de richting biologie. Desondanks zijn jullie mij altijd onvoorwaardelijk blijven steunen.
Aan Oscar, de allerbeste kat ooit… Vanaf de eerste examens van het lager tot de laatste examens van het hoger. Dju, net niet het laatste obstakel. Kameraad, bedankt voor al die jaren trouw aan mijn zijde.
2
Inhoud
Dankwoord ... 1
Inleiding ... 5
Probleemstelling ... 5
De kleine marterachtigen: de centrale rol van lichaamsgrootte ... 7
De wezel – Mustela nivalis ... 8
De hermelijn – Mustela erminea ... 11
Bedreigingen in Vlaanderen: een risicoanalyse ... 14
Skrjabingylus nasicola ... 15
Monitoring en citizen science ... 17
Doelstellingen ... 19
Autopsiegedeelte ... 19
Doodsoorzaak en conditieparameters ... 19
Variatie in vachtkarakteristieken ... 19
Leeftijdscohorten ... 20
Lichaamsgrootte en seksueel dimorfisme ... 20
Reproductie ... 20
S. nasicola ... 20
Dieet ... 20
Pootafmetingen wezel en hermelijn ... 20
Toekomstig onderzoek... 20 Citizen science ... 21 Materiaal en methoden ... 22 Autopsiegedeelte: ... 22 INBO-protocol ... 22 Algemene situering ... 22
Doodsoorzaken in relatie met conditieparameters ... 22
Variatie in vachtkarakteristieken ... 22
Leeftijdscohorten ... 24
Lichaamsgrootte en seksueel dimorfisme ... 25
Reproductie ... 27
S. nasicola ... 28
3
Maaganalyse kleine marterachtigen ... 29
Pootafmetingen wezel en hermelijn ... 31
Citizen science ... 33 Dataexploratie ... 33 Populatietrend ... 33 Resultaten ... 35 Autopsiegedeelte ... 35 Wezel ... 35 Hermelijn ... 48 Wezel en hermelijn ... 62
Maaganalyse van de kleine marterachtigen (huidig onderzoek en Verschuren (2005)). ... 62
Poten: mannetjes wezel & vrouwtjes hermelijn ... 72
Monitoring en citizen science ... 78
Dataexploratie ... 78
Populatietrend ... 80
Discussie ... 84
Autopsiegedeelte ... 84
Doodsoorzaken in relatie met conditionele parameters ... 84
Wintervacht hermelijn ... 84
Lichaamsafmetingen ... 85
Reproductie ... 86
Skrjabingylus nasicola ... 89
Maaganalyse van de kleine marterachtigen ... 90
Pootafmetingen wezel en hermelijn ... 92
Monitoring en citizen science ... 92
Conclusie ... 94
Samenvatting ... 96
Appendix ... 98
Opbouw vacht en taxonomische kenmerken ... 98
DSB-methode en leeftijd gerelateerde ontwikkelingen van schedel en baculum... 100
He04/011: winterreproductie? ... 102
Inktafdrukken poten ... 103
4 Literatuurlijst ... 109
5
Inleiding
Probleemstelling
De inheemse carnivorengemeenschap van Vlaanderen kende de laatste decennia enkele opmerkzame veranderingen met voornamelijk de middelgrote carnivoren die op heden een grotere verspreiding laten noteren. In de eerste plaats evolueerde de vos (Vulpes vulpes) reeds voor de eeuwwisseling tot een algemene soort (Van Den Berge, 1995) gevolgd door de steenmarter (Martes fiona) (Van Den Berge et al., 2017). Daarnaast worden zowel de das (Meles meles) als de boommarter (Martes martes) steeds frequenter geobserveerd in alle Vlaamse provincies (Van Den Berge et al., 2015; Van Den Berge et al., 2017). Een omgekeerde beweging lijkt zich in te zetten voor de kleine marterachtigen. In het geval van de bunzing (Mustela putorius) wordt een duidelijke afname in het aantal verkeersslachtoffers gemeld wat een achteruitgang doet vermoeden (Van Den Berge & Gouwy, 2012). Ook omtrent de wezel (Mustela
nivalis) en de hermelijn (Mustela erminea), de kleinste vertegenwoordigers van de Vlaamse
carnivorengemeenschap, heerst reeds enkele decennia het idee dat beide soorten minder zouden voorkomen (Van Den Berge & De Pauw, 2003).
Van zowel de wezel als de hermelijn wordt aangenomen dat beide carnivoren een gebiedsdekkende verspreiding zouden vertonen in Vlaanderen. Dit is grotendeels gebaseerd op vangstresultaten gepubliceerd in oude jachttijdschriften (Van Den Berge & De Pauw, 2003). Sinds het stopzetten van deze praktijken is omtrent de actuele verspreiding echter weinig betrouwbare informatie beschikbaar. De verborgen levenswijze en van nature lage populatiedichtheden verhinderen immers het regelmatig waarnemen van beide marterachtigen. In een ideaal scenario zouden de wezel en de hermelijn gemonitord worden aan de hand van indirecte waarnemingen volgens een gestandaardiseerd telprotocol. Dergelijk onderzoek is echter in de eerste plaats niet tot amper toepasbaar op landschapsschaal en/of op lange termijn omwille van een gebrek aan middelen. Ten tweede vereist de kleine lichaamsgrootte van de wezel en de hermelijn een minimum aan ervaring met betrekking tot sporenonderzoek, zeker in afwezigheid van een jaarlijks sneeuwlandschap, of alternatieve monitoringstechnieken.
Het monitoren van carnivoren vormt in het algemeen een grote uitdaging en als reactie hierop richtte het Instituut voor Natuur- en Bosonderzoek (INBO) in 1998 het Marternetwerk op. Dit initiatief, dat beroep doet op vrijwilligers, richt zich tot het samenbrengen van verspreidingsgegevens van inheemse en uitheemse carnivoren op basis van verkeersslachtoffers en de inzameling hiervan. Hoewel het project zijn nut bewijst, bijvoorbeeld in het geval van de bunzing en de steenmarter (Van Den Berge & Gouwy, 2012; Van Den Berge et al., 2012), blijven de wezel en de hermelijn duidelijk onder de radar van dit project. Door de geringe lichaamsgrootte vallen verkeersslachtoffers immers amper op of worden deze snel van de rijbaan verwijderd door aaseters, zoals bijvoorbeeld kraaiachtigen. Slechts een beperkte hoeveelheid exemplaren wordt bijgevolg op jaarbasis ingezameld. Een aantal dat niet toereikend is om een inkijk te bieden op de hedendaagse verspreiding of populatieontwikkeling (Van Den Berge and De Pauw, 2003). In 2014 is de geüpdatete versie van de IUCN Rode Lijst van de Zoogdieren in Vlaanderen gepubliceerd en deze vormt een eerste samenvattende bron met betrekking tot een verandering in verspreiding van beide kleine marterachtigen. Op basis van het verschil in vastgestelde aanwezigheden binnen onderzochte kilometerhokken tussen de periodes 1987-2002 en 2003-2013, wordt een achteruitgang voor beide soorten vastgesteld. Bijgevolg wordt de wezel volgens de regionale criteria toegewezen tot de conservatiestatus ‘Bijna in gevaar’. Voor de hermelijn wordt naast de aangetoonde daling in verspreiding bovendien het aantal zich voortplantende exemplaren op minder dan 10.000 geschat. Om deze reden
6 wordt het risico op regionaal uitsterven een categorie hoger bepaald, wat zich vertaalt in de conservatiestatus ‘Kwetsbaar’ (Maes et al., 2014).
Globaal wordt de conservatiestatus van beide kleine marterachtigen aangeduid als ‘Niet bedreigd’ uitgaande van de wijde holarctische verspreiding en de veronderstelde stabiele populatiegroottes (McDonald et al., 2016; Helgen & Kranz, 2016). Toch heerst ook buiten Vlaanderen het idee dat in meerdere antropogeen gestuurde regio’s binnen Europa de wezel en de hermelijn een neerwaartse populatietrend zouden ervaren. Vaak wordt hierbij de aangaande fragmentatie van het extensieve cultuurlandschap als mogelijke oorzaak aangeduid (Müri, 2009; Graf et al., 2014; Van Maanen et al., 2015). In Nederland wordt dit vermoeden overigens bevestigd door de regionale Rode Lijst. Beide soorten worden er toegewezen aan de categorie ‘Bijna in gevaar’ op basis van een lagere vastgestelde aanwezigheid binnen onderzochte kilometerhokken (Zoogdiervereniging, 2006). De conservatiestatus in de overige buurregio’s en/of -landen duidt beide soorten aan als niet bedreigd, is niet recent beoordeeld of is niet bepaald omwille van een onvoldoende hoeveelheid data (tabel 1).
Regio/land Wezel (M. nivalis) Hermelijn (M. erminea) Referenties Vlaanderen Bijna in gevaar Kwetsbaar Maes et al. (2014) Wallonië Momenteel niet in gevaar Momenteel niet in gevaar Libois (1997)
Nederland Bijna in gevaar Bijna in gevaar Zoogdiervereniging (2006) Frankrijk: Momenteel niet in gevaar Momenteel niet in gevaar IUCN France (2017) Duitsland: Onvoldoende data Onvoldoende data Meinig et al. 2009
Noordrijn-Westfalen Onvoldoende data Onvoldoende data Meinig et al. 2011
Tabel 1: de regionale conservatiestatus van de wezel en de hermelijn in Vlaanderen en zijn buurregio’s -landen.
Hoewel op basis van de regionale IUCN-criteria per definitie de kans op regionaal uitsterven reëel wordt geacht (Maes et al., 2014), is naast de actuele verspreiding ook weinig informatie beschikbaar omtrent de ecologie van beide soorten in Vlaanderen. Deze kennis is nodig in functie van het identificeren van doelgerichte conservatiemaatregelen. Een initiële aanzet tot het verzamelen van soortspecifieke kennis vond plaats in het kader van een scriptie uitgevoerd door Verschuren (2005). Tijdens dit onderzoek werd een eerste reeks autopsies verricht op karkassen van beide soorten die werden ingezameld door het Marternetwerk. Onderhavige thesis heeft tot doel navolging te geven aan dit werk door een tweede reeks autopsies te verrichten en de mogelijkheden van citizen science met betrekking tot monitoring te exploreren. In het vervolg van dit hoofdstuk worden beide soorten kort geïntroduceerd aan de hand van de wetenschappelijke literatuur met een focus op morfologische variatie, voedselecologie en reproductie. Hierbij wordt teruggekoppeld naar de resultaten van Verschuren (2005). Ook worden mogelijke bedreigingsfactoren in verband met het voortbestaan van beide marterachtigen in Vlaanderen aangesneden waarna summier de mogelijkheden van alternatieve monitoringstechnieken en citizen science worden voorgesteld.
7
De kleine marterachtigen: de centrale rol van lichaamsgrootte
De wezel en de hermelijn behoren tot de marterachtigen en nemen binnen deze ecologisch diverse familie een duidelijke positie in. Het zijn solitaire, kortlevende en gespecialiseerde carnivoren die zich voornamelijk voeden met kleine knaagdieren (King & Moors, 1979; Macdonald et al., 2017). Deze specialisatie vloeit voort uit een uitgesproken morfologie, namelijk een dun en langgerekt lichaam met korte poten en een geringe lichaamsgrootte. Deze eigenschappen laten toe kleine knaagdieren doeltreffend te bejagen tot in sterk besloten ruimtes zoals ondergrondse holen en gangenstelsels (King & Powell, 2006). De wezel en de hermelijn worden hierbij geholpen door een korte vacht en een laag vetpercentage (Casey & Casey, 1979; King, 1989; Zielinski, 2000).
Beide kleine marterachtigen worden gekenmerkt door seksueel dimorfisme waarbij het mannelijke geslacht beduidend groter is (King and Powell, 2006). Een oorspronkelijke verklaring berust op het verdelen van voedselbronnen zodanig interseksuele competitie zich tot een minimum zou beperken (Erlinge, 1979; Dayan & Simberloff, 1994). Een meer geaccepteerde hypothese duidt daarentegen seksuele selectie aan als oorzaak. Hierbij is een grotere lichaamsgrootte een voordeel voor het mannelijke geslacht in de onderlinge concurrentiestrijd tot copulatie in een polygynous paringssysteem. Het vrouwelijke geslacht maximaliseert daarentegen de foerageerefficiëntie dankzij een kleinere lichaamsgrootte en bijgevolg, het succesvol grootbrengen van jongen (Simms, 1979; King, 1989).
Hoewel uiterst geschikt tot het bejagen van kleine knaagdieren, impliceert het bouwplan van de wezel en de hermelijn ook meerdere nadelen. De oppervlakte/volume-verhouding, de korte vacht en een laag vetpercentage staan synoniem voor een snel metabolisme dat het frequent nuttigen van energierijke maaltijden vereist (Brown & Lasiewski, 1972; King, 1989; Zielinski, 2000). Daarnaast zijn algemeen slechts weinig alternatieve prooien beschikbaar die op een efficiënte wijze deze energetische noden kunnen vervullen (Zielinski, 2000; King & Powell, 2006). Bijgevolg wordt een duidelijke relatie genoteerd tussen de populatiedichtheid van beide kleine marterachtigen en die van hun primaire prooidiersoorten (Debrot, 1981; McDonald et al., 2000; Hanski et al., 2001). In extreme, maar niet uitzonderlijke gevallen, treedt hierbij zelfs het lokaal verdwijnen op (King, 1989; King & Powell, 2006). Beide soorten worden echter ook gekenmerkt door een hoge reproductiecapaciteit indien een groot prooiaanbod aanwezig is (King & Moors, 1979), en daarnaast beschikken ze tevens over een sterk dispersievermogen (Burns, 1964; Murphy & Dowding, 1994; Murphy & Dowding, 1995).
Als kleine predatoren zijn de wezel en de hermelijn gevoelig voor interferentiecompetitie met enerzijds middelgrote carnivoren en anderzijds dag- en nachtroofvogels (Zielinski, 2000). Beide soorten worden op een regelmatige basis aangetroffen in het dieet van deze grotere competitoren (King & Powell, 2006; Dell'Arte et al. 2007; Palazón et al., 2016). De aanwezigheid van interferentiecompetitie heeft een invloed op de nichedimensie van de kleine marterachtigen en habitatstudies duiden onder meer op het belang van dekkinggevende structuren in het landschap ter bescherming (Klemola et al., 1999; Zub et al., 2008). De precieze impact van grotere predatoren op de populatiedynamiek is daarentegen niet gekend. Wel wordt in één geval het lokaal verdwijnen van de hermelijn in het duinlandschap tussen Den Haag en Den Helder in verband gebracht met de toenmalige rekolonisatie van de vos (Mulder, 1990).
Niettegenstaande beide soorten op elkaar lijken, zowel qua gedaante als algemene ecologie, verschillen de wezel en de hermelijn met betrekking tot dieet, reproductie en habitatvoorkeur met een centrale rol weggelegd voor lichaamsgrootte. Deze verschillen vormen de basis voor een samenspel van competitieve voordelen die coëxistentie op lokale en regionale schaal mogelijk maken (King & Moors, 1979).
8
De wezel
– Mustela nivalis
De wezel heeft een holarctische verspreiding (McDonald et al., 2016) en is in Europa vrijwel overal aan te treffen met uitzondering van Ierland, bepaalde regio’s in de Balkan en het binnenland van Spanje (Van Den Berge & De Pauw, 2003). Het areaal van de soort is door menselijk toedoen verder uitgebreid tot onder meer Nieuw-Zeeland en verschillende eilanden van het Middellandse Zeegebied onder de noemer van biologische controle. Voorts wordt ook de aanwezigheid in Noordwest-Afrika in verband gebracht met herhaalde introducties door de mens. (McDonald
et al., 2016).
De wezel onderscheidt zich van de hermelijn door een proportioneel kortere staart zonder zwarte pluim en een geringere lichaamsgrootte waar beide soorten sympatrisch voorkomen (Macdonald et al., 2017). De wezel wordt tevens aangeduid als de kleinste
vertegenwoordiger van de gehele carnivorenorde, hoewel deze titel voornamelijk van toepassing is voor populaties die Noord-Amerika en het noordelijke verspreidingsgebied van Eurazië bevolken (King & Powell, 2006). Lichaamsgrootte vertoont immers een grote mate van variatie doorheen het areaal en het algemeen patroon wijst op een grotere lichaamsgrootte waar de primaire prooidiersoorten omvangrijker zijn (Erlinge, 1987; King, 1991b). Binnen Europa wordt voornamelijk, in tegenstelling tot de regel van Bergmann, een kleinere lichaamsgrootte geobserveerd met een toenemende latitudinale gradiënt (Rensch, 1938; King & Powell, 2006; Yom-Tov et al., 2010) (tabel 2).
Kop-romplengte (mm) Mannetjes, adult referenties
Sint-Petersburg 177 (n=22) Danilov & Tumanov (1976); Abramov & Baryshnikov (2000) Engeland 202 (n=46) Corbet & Harris (1991)
Duitsland 181 (n=138) Reichstein (1993)
Centraal-Frankrijk 218 (n=46) Abramov & Baryshnikov (2000) Bohemen/Moravië 217 (n=355) Abramov & Baryshnikov (2000) Spanje 244 (n=6) Miller (1912); Cabrera (1914) Griekenland 336 (n=28) Douma-Petridou & Ondrias (1987)
Vlaanderen 196 (n=81) Verschuren (2005)
Tabel 2: de gemiddelde kopromplengte van de adulte mannelijke wezel in verschillende Europese regio’s met de resultaten van
Verschuren (2005) hieraan toegevoegd.
Doorheen het areaal wordt bovendien ook een opmerkelijke mate van variatie vastgesteld met betrekking tot onder meer het kleurpatroon van de (winter)vacht, de mate van seksueel dimorfisme en de relatieve lengte van de staart (Abramov & Baryshnikov, 2000; King & Powell, 2006). Op basis van de morfologische variatie onderscheiden Abramov & Baryshnikov (2000) 19 ondersoorten.
Figuur 1: morfologische variatie van de wezel met
bovenaan de ondersoort vulgaris met een grillige demarcatielijn en onderaan de ondersoort nivalis met een rechte demarcatielijn en afwezigheid van bruine vlekken.
9 Omwille van conventionele redenen worden meestal slechts twee ondersoorten vermeld in de literatuur (King & Powell, 2006; Macdonald et al., 2017):
• Mustela nivalis vulgaris: West- en Zuid Europa en Noordwest-Afrika (Macdonald et al., 2017) • Mustela nivalis nivalis: Noord-Europa, Azië en Noord-Amerika (Macdonald et al., 2017)
De ondersoort aanwezig in Vlaanderen, namelijk Mustela nivalis vulgaris, wordt gekenmerkt door een grillige demarcatielijn tussen de dorsaal vaalbruine en de ventraal witgekleurde vacht (Van Den Berge & De Pauw, 2003). Voorts kunnen nabij de demarcatielijn bruine vlekken van verschillende grootte en vorm in de witte vacht worden aangetroffen over de volledige lengte van het lichaam. Ter hoogte van de mondhoeken is langs beide zijden een bruine wangvlek aanwezig die in bepaalde gevallen kan overgaan met de dorsaal bruingekleurde vacht. De wintervacht kleurt niet wit maar behoudt het kleurpatroon van de zomervacht (Abramov & Baryshnikov, 2000). De ondersoort Mustela nivalis nivalis wordt daarentegen gekenmerkt door een rechte demarcatielijn en de afwezigheid van bruine (wang)vlekken. Mustela nivalis
nivalis kleurt wel wit gedurende de winterperiode (Frank, 1985; Abramov & Baryshnikov, 2000) (figuur 1).
De resultaten van Verschuren (2005) vertonen hieromtrent een inconsistentie door het vermelden van de aanwezigheid van een rechte demarcatielijn. In deze dataset noteert de auteur een rechte lijn in 14.5% (n=56) van de geautopseerde wezels. Daarnaast wordt ook variatie vastgesteld met betrekking tot het kleurpatroon van de poten. In ongeveer de helft (56.1%, n=65) van de onderzochte wezels kleuren de tenen van de voorpoten wit en bij een klein aantal zijn de tenen van de achterpoten eveneens wit gekleurd (15.9%, n=65).
De wezel wordt beschreven als een uiterst gespecialiseerde predator van Microtus woelmuizen (Day, 1966; Erlinge, 1975; Moors, 1975; Korpimäki et al., 1991). Afhankelijk van het plaatselijke en temporele prooiaanbod worden ook grote aantallen bosmuizen (Apodemus spp.) en woelmuizen van het genus
Myodes geconsumeerd (Erlinge, 1975; Tapper, 1979; Jędrzejewski et al., 1992; Jedrzejewski et al., 1995;
McDonald et al., 2000; Elmeros, 2006). Secundaire prooidiercategorieën van de wezel zijn insectivoren (Insectivora), (jonge) haasachtigen (Lagomorpha), vogels en diens eieren (Aves) (Erlinge, 1975; Brugge, 1977; Elmeros, 2006). De relatieve inbreng van deze categorieën neemt typisch toe onder voedselarme condities maar blijft eerder beperkt (King & Powell, 2006). Voorts worden sporadisch ook kikkers, hagedissen en ongewervelden teruggevonden in het dieet (Elmeros, 2006; Lanszki & Heltai, 2007). Het seksueel dimorfisme kan zich uiten door een breder gebruik van het voedselspectrum bij het mannelijke geslacht (Erlinge, 1975; Tapper, 1979) maar wordt niet consistent waargenomen in de literatuur (McDonald et al., 2000).
Binnen de carnivorenorde verhoudt de wezel zich als een uitgesproken r-strateeg omwille van een groot aantal jongen, een vroege seksuele rijpheid en de mogelijkheid tot twee worpen per jaar (King & Moors, 1979). Spermatogenese en oestrus vatten aan in februari en vanaf maart worden de eerste zwangerschappen in het wild geobserveerd. De draagtijd duurt 35 tot 37 dagen en de eerste geboortes worden vastgesteld in april. Een volledige reproductiecyclus, inclusief het grootbrengen van de jongen, neemt drie tot vier maanden in beslag (King & Powell, 2006). De gemiddelde worpgrootte bedraagt 5-6 jongen maar dit aantal kan sterk variëren (East & Lockie, 1964; King & Moors, 1979; Sundell, 2003; King & Powell, 2006). In de dataset van Verschuren (2005) werden twee drachtige vrouwtjes vastgesteld. Het ene vrouwtje telde drie embryo’s en het andere acht.
10 Mannetjes blijven vruchtbaar tot minimum augustus en negen tot tien weken na het werpen van de jongen komen de adulte vrouwtjes opnieuw in oestrus, waarna een tweede reproductiecyclus kan starten. De geboortes van deze tweede cyclus concentreren zich in juli en augustus (Frank, 1974; King & Powell, 2006). In de dataset van McDonald & Harris (2002) (n=77) werd een laatste dracht vastgesteld op 5 oktober. Daarnaast bereiken de eerste jongen van het jaar seksuele volwassenheid na drie tot vier maanden en deze kunnen op hun beurt overgaan tot reproductie vanaf juli (King & Powell, 2006). Ondanks de mogelijkheid tot reproductie later op het jaar situeert het hoogtepunt van het voortplantingsseizoen zich voornamelijk in de lenteperiode (Van Den Berge & De Pauw, 2003). Een tweede worp is namelijk enkel succesvol indien rijke voedselcondities van toepassing zijn en onder een zeer laag prooiaanbod mislukt zelfs elke poging, inclusief de eerste (King & Powell, 2006). Jedrzejewski et al. (1995) toonden hierbij aan dat een minimum van 10 tot 14 woelmuizen per hectare vereist is om over te gaan tot succesvolle voortplanting.
Verschuren (2005) bevestigt in haar dataset grotendeels de in de literatuur vermeldde voortplantingsperiode door de aanwezigheid van vrij sperma aan te tonen vanaf februari tot en met augustus, en één uitzondering in december. Hierbij werd niet nagegaan of ook eerstejaars wezels reproductief zijn. Daarnaast wordt slechts een fragmentarisch beeld gegeven omtrent de reproductiecyclus bij vrouwtjes te wijten aan het lage aantal vrouwelijke exemplaren in de dataset. Enkel in juni en juli werden (recent) drachtige vrouwtjes vastgesteld.
Vrouwtjes bereiken fysieke volwassenheid op een leeftijd van 3-4 maanden en bij mannetjes is dit op een leeftijd van 3-6 maanden. Het zijn voornamelijk de jongen die later op het jaar worden geboren, die een tragere ontwikkeling zouden hebben (King & Powell, 2006). De wezel kan een leeftijd van vijf tot zes jaar bereiken, maar wordt gemiddeld niet ouder dan één jaar (King & Moors, 1979; Van Den Berge & De Pauw, 2003).
Doorheen het verspreidingsgebied wordt aanwezigheid in een grote variëteit van habitattypes vastgesteld. De soort wordt naast het extensieve cultuurlandschap onder meer aangetroffen in duin- en bosgebieden tot zelfs tuinen en akkerlandcomplexen (Klemola et al., 1999; Van Den Berge & De Pauw, 2003; Macdonald et al., 2004; Zub et al., 2008; Magrini et al., 2009; McDonald et al., 2016). De wezel lijkt hierbij niet zozeer aan een welbepaalde biotoop op landschapsschaal gebonden te zijn, maar is voornamelijk afhankelijk van aanwezige dekkinggevende (micro)structuren (Van Den Berge & De Pauw, 2003). Deze relatie wordt in grote mate weerspiegeld door het habitatgebruik waarbij zelden wordt afgeweken van lijnvormige elementen (Erlinge, 1974; Macdonald et al., 2004; Wilson & Reeder, 2005; Schmitt, 2006). Het belang van deze structuren wordt enerzijds verklaard door hogere dichtheden van prooien (Klemola et al., 1999) en anderzijds dienen ze, zoals reeds vermeld, ter bescherming tegen grotere predatoren (Klemola et al., 1999; Zub et al., 2008). Naast het algemeen vermijden van open zones wordt ook een negatieve selectie geobserveerd voor vochtige gebieden. Sidorovich et al. (2008) verklaren dit door interferentiecompetitie met de hermelijn (zie verder).
11 Figuur 2: verspreidingsgebied van de wezel (Mustela nivalis) op de noordelijke hemisfeer (McDonald et al., 2016).
De hermelijn
– Mustela erminea
De verspreiding van de hermelijn omvat grote delen van het holarctische gebied (Helgen & Kranz, 2016) maar reikt in tegenstelling tot de wezel minder zuidelijk in Eurazië (figuur 4). Op het Europese grondgebied vertaalt zich dat in een afwezigheid in het mediterrane gebied en de zuidelijke en oostelijke regio van de Balkan (Van Den Berge & De Pauw, 2003). De soort is daarentegen wel aanwezig in Ierland. Net als de wezel is de hermelijn vanuit Groot-Brittannië geïntroduceerd in Nieuw-Zeeland waar hij uitgroeide tot een invasie exoot. Tot op heden teistert de soort er nog steeds het inheemse vogelbestand ondanks doorgedreven controlemaatregelen (King, 2019). Doorheen het verspreidingsgebied worden 37 ondersoorten erkend (Wilson & Reeder, 2005). De ondersoort aanwezig in West- en Centraal-Europa en Europees Rusland wordt aangeduid als Mustela
erminea aestiva (Heptner & Sludskii, 2002).
Doorheen het verspreidingsgebied vertoont de hermelijn een grote mate van variatie in lichaamsgrootte (King & Powell, 2006) en analoog met de wezel wordt op het Europese continent een geringere grootte richting het noorden vastgesteld (Yom-Tov et al., 2010) Een opvallende uitzondering is echter Groot-Brittannië waar veruit de grootste exemplaren van Europa worden geobserveerd (Corbet & Harris, 1991) (tabel 3). Net als bij de wezel wijst het algemeen patroon op een toenemende lichaamsgrootte waar de primaire prooidiersoorten omvangrijker zijn (Erlinge, 1987; King, 1991a).
Figuur 3: De hermelijn in volledig witte wintervacht op de
12 Totaal gewicht (gram) Man, adult Vrouw, adult Referenties
Noord-Zweden 184 (n=19) 98 (n=6) Erlinge (1987) Zuid-Zweden 230 (n=73) 137 (n=25) Erlinge (1987)
Groot-Brittannië 321 (n=204) 213 (n=99) Corbet & Harris (1991) Zwitserland 46-47°N 241 (n=72) 157 (n=43) Debrot (1981)
Frankrijk 46-47°N 243 (n=21) 145 (n=6) Delattre, niet gepubliceerd
Vlaanderen 255 (n=45) 169 (n=16) Verschuren (2005)
Tabel 3: totaal gewicht van de hermelijn in Europa (King & Powell, 2006) met de resultaten van Verschuren (2005) toegevoegd.
Naast lichaamsgrootte onderscheidt de hermelijn zich van de wezel door een proportioneel langere staart met een zwarte kwast. De zomervacht is tweekleurig met een grijs- tot vaalbruine rugzijde en een witte tot geelwitte buikzijde. De demarcatielijn tussen beide kleuren is recht (Van Den Berge & De Pauw, 2003) (figuur 4), hoewel in Noord-Amerika en bepaalde populaties in Ierland een golvende demarcatielijn wordt waargenomen (King & Powell, 2006). Met betrekking tot de witte of geelwitte buikzijde noteert Verschuren (2005) in 60.3% van de geautopseerde hermelijnen de geelwitte variant (n=58).
Afhankelijk van de klimatologische regio kleurt de wintervacht volledig wit, op de zwarte staartpunt na, of wordt het patroon van de zomervacht behouden. In de transitiezone wordt daarnaast ook een gevlekt fenotype waargenomen (King & Powell, 2006). In Vlaanderen worden vooralsnog alle drie de vormen genoteerd maar de onderlinge verdeling hiervan is niet gekend (Van Den Berge & De Pauw, 2003; Verschuren, 2005). De start van de rui wordt gedreven door (in)activatie van de hypofyse met betrekking tot melatonineproductie onder invloed van de hoeveelheid daglicht (Zimova et al., 2018). Gebaseerd op de dataset van Van Soest & Van Bree (1969) wordt in Nederland de aanvang van het verruilen van de zomervacht voor de wintervacht ten vroegste vastgesteld begin november. Omgekeerd, verdwijnen volledig witte exemplaren ten laatste eind maart uit beeld. Daarnaast observeerden Van Soest & Van Bree (1969) een duidelijk groter percentueel aantal witte en gevlekte fenotypes in Nederlandse provincies met een gemiddeld koudere temperatuur in januari.
In termen van exploitatiecompetitie wordt de hermelijn beschreven als de minder efficiënte predator in vergelijking met de wezel. De meer omvangrijke lichaamsgrootte impliceert dat de soort in mindere mate toegang heeft tot de meer besloten ruimtes (Simms, 1979; Pounds, 1981; Erlinge & Sandell, 1988). Dit wordt weerspiegeld door een gemiddeld hogere consumptie van grotere prooidiersoorten en een breder gebruik van het voedselspectrum (Brugge, 1977; McDonald et al., 2000; Elmeros, 2006). Hoewel woelmuizen van het genus Microtus op het Europese continent steeds een duidelijk aandeel van het dieet vormen (King & Powell, 2006), wordt het belang van de grotere woelrat (Arvicola terrestris) aangeduid in verschillende regio’s. Een duidelijke associatie tussen de populatiedynamiek van de hermelijn en de densiteit van woelratten vindt men terug in Europees-Rusland, Wit-Rusland, Zuid-Zweden en Zwitserland (Aspisov & Popov, 1980; Debrot, 1981; Sidorovich & Solovej, 2007). De crash van de hermelijnenpopulatie in Groot-Brittannië tijdens de myxomatose uitbraak gedurende de jaren ’50, ’60 en ’70 van de vorige eeuw, is daarentegen getuige van een specialisatie op konijnen (Oryctolagus cuniculus) (Day, 1966; McDonald et
al., 2000). Op het Europese vasteland, waar de hermelijn een kleinere lichaamsgrootte heeft, kunnen
haasachtigen eerder worden beschouwd als een secundaire prooidiercategorie, hoewel dit een hoge consumptie in tijd en ruimte (bv. in een duinlandschap of tijdens de lente) niet uitsluit (Brugge, 1977; Debrot, 1981; Erlinge, 1981). Ook vormen vogels en diens eieren een belangrijke, zo niet de belangrijkste, alternatieve voedselbron (Brugge, 1977; Piontek et al., 2015). Voorts wordt het dieet binnen Europa, afhankelijk van habitat en tijd, verder aangevuld met Myodes woelmuizen en bosmuizen en in mindere
13 mate met spitsmuizen, hagedissen en amfibieën (Erlinge, 1981; Jedrzejewska & Jedrzejewski, 2013; Piontek et al., 2015, Macdonald et al., 2017). In geïsoleerde gevallen wordt ook consumptie van de jeneverbes (Juniperus communis) en aas vastgesteld (Martinoli et al. 2001; Piontek et al. 2015). Het seksueel dimorfisme kan zich net zoals bij de wezel uiten door een breder gebruik van het voedselspectrum en consumptie van gemiddeld grotere prooisoorten door mannetjes (Erlinge, 1981; McDonald et al. 2000), maar ook voor de hermelijn wordt deze relatie niet consistent aangetoond (Piontek et al., 2015).
Desalniettemin een duidelijke relatie wordt aangetoond tussen de populatiedynamiek van de hermelijn en diens primaire prooidiersoorten (Aspisov & Popov, 1980; Debrot, 1981; McDonald et al, 2000; Sidorovich & Solovej, 2007), wordt op basis van een breder voedselspectrum aangenomen dat de soort perioden van voedselschaarste beter weerstaat in vergelijking met de wezel (King & Powell, 2006). Deze assumptie wordt weerspiegeld in de voortplantingsecologie van de hermelijn die slechts één reproductiecyclus per jaar telt.
De reproductie-ecologie van de hermelijn wordt hierbij gekenmerkt door uitgestelde implantatie wat betekent dat eicellen na bevruchting wel ontwikkelen tot blastocysten maar vervolgens in kiemrust gaan. Gedurende meerdere maanden blijven de blastocysten vrij zweven in de uterus ten gevolge van de secretie van een lage hoeveelheid progesteron geproduceerd door de corpora lutea of gele lichaampjes. Als reactie op de toename van de hoeveelheid daglicht na de winterperiode, produceert de hypofyse het luteïniserend hormoon waarna de productie van progesteron verhoogt en de implantatie van de blastocysten enkele dagen later plaatsvindt. Na de implantatie volgt 28 dagen later de geboorte die temporeel sterk gesynchroniseerd is binnen een populatie (King & Powell, 2006). Wanneer deze juist plaatsvindt is afhankelijk van de klimatologische regio en wordt voor Vlaanderen door Van Den Berge & De Pauw (2003) rond april-mei gesitueerd. De gemiddelde worpgrootte situeert zich tussen zes en negen jongen, afhankelijk van de auteur, en kan bovendien sterk oplopen (King & Moors, 1979; Ternovsky, 1983). Verschuren (2005) vermeldt in haar dataset twee zwangere vrouwtjes, ingezameld begin april (6/4 en 9/4), waarvan de eerste acht embryo’s telde en de tweede 10.
De spermatogenese bij mannetjes wordt eveneens geactiveerd als reactie op een toenemende hoeveelheid zonlicht. Vrij sperma in de epidydimi wordt vanaf maart geobseerveerd door Verschuren (2005). Adulte vrouwtjes komen terug in oestrus nog tijdens het zogen en zijn bijgevolg klaar voor copulatie (King & Moors, 1979). Daarnaast zijn juveniele vrouwtjes seksueel volwassen nog voor het einde van de zoogperiode en zijn op hun beurt klaar om bevrucht te worden, nog voor het nest verlaten wordt (King & Moors, 1979; Van Den Berge & De Pauw, 2003). Juveniele vrouwtjes bereiken uiteindelijk fysieke volwassenheid op een leeftijd van zes maanden. Het groeiproces van juveniele mannetjes wordt daarentegen onderbroken na een initiële groeispurt en pas op een leeftijd van ongeveer 12 maanden wordt fysieke en seksuele volwassenheid bereikt (King & Powell, 2006). De maximumleeftijd van de hermelijn is 10 jaar, maar de werkelijke leeftijdsverwachting ligt waarschijnlijk veel lager (Van Den Berge & De Pauw, 2003).
De hermelijn doorkruist doorheen zijn verspreidingsgebied een brede range van habitattypes (Macdonald
et al., 2017). De soort wordt onder meer aangetroffen langsheen rivieroevers en bosranden, in duin-,
moeras- en veengebieden en landschappen met een intense afwisseling tussen grasland en lijnvormige, dekkinggevende structuren (Debrot & Mermod, 1983; Klemola et al., 1999; Van Den Berge & De Pauw, 2003; Hellstedt & Henttonen, 2006; Vogel, 2006; Sidorovich et al., 2008; Červinka et al., 2013). De literatuur duidt hierbij op een voorkeur voor vochtigere gebieden dat wordt gerelateerd aan de
14 aanwezigheid van grotere knaagdieren zoals de woelrat (Debrot, 1981; Sidorovich & Solovej, 2007). In Groot-Brittannië wordt de associatie gelegd met de aanwezigheid van het konijn los van een vochtig habitattype (King & Moors, 1979). Voorts lijken grote continue boscomplexen vermeden te worden hoewel dit aanwezigheid niet uitsluit (Debrot & Mermod, 1983; Bounous et al., 1995; Hellstedt & Henttonen, 2006).
Figuur 4: verspreidingsgebied van de hermelijn op de noordelijke hemisfeer (Helgen & Kranz, 2016).
Bedreigingen in Vlaanderen: een risicoanalyse
Het prooiaanbod vormt de belangrijkste verklarende variabele omtrent de aan- of afwezigheid van beide gespecialiseerde soorten. Meer specifiek wordt in de literatuur verwezen naar het belang van de Microtus woelmuizen en de woelrat op de populatiedynamiek van de wezel en de hermelijn (Debrot, 1981; McDonald et al., 2000; Hanski et al., 2001). De IUCN Rode Lijst van de Zoogdieren in Vlaanderen duidt echter alle leden van het Microtus genus en de woelrat aan tot de categorie ‘Bijna in gevaar’. Daarnaast behoren ook de haasachtigen, waarbij meer bepaald het wild konijn (Oryctolagus cuniculus) mogelijk een belangrijke secundaire prooidiersoort is in Vlaanderen, tot deze conservatiecategorie (Maes et al., 2014). De bedreigde status van beide kleine marterachtigen (en die van hun prooidiersoorten) wordt onder meer in Nederland en Zwitserland toegeschreven aan de huidige toestand van het extensieve cultuurlandschap (Müri, 2009; Van Maanen et al., 2015). Ook in Vlaanderen is dit landschap in verval sinds de tweede helft van de 20e eeuw ten gevolge van een intensifiëring van de landbouw en doorgedreven urbanisatie (Maes
& Van Dyck, 2001). Habitatfragmentatie wordt aangeduid als één van de belangrijkste oorzaken voor de huidige biodiversiteitsproblematiek (Turner, 1996): het leidt enerzijds tot een reductie van de totale habitatoppervlakte en -kwaliteit, en creëert anderzijds geïsoleerde leefgebieden binnenin een onherbergzame matrix (Frankham et al. 2010). In de overgebleven kleinere habitatpatches zijn populatiegroottes kleiner, resulterend in een hogere gevoeligheid voor lokale extinctie ten gevolge van demografisch-, genetisch- en omgevingsgerelateerde stochastische effecten (Young et al., 1996; Bender
et al., 1998; Fahrig, 2003). Daarbij wordt succesvolle dispersie in toenemende mate verhinderd waardoor
deze effecten mogelijk worden geamplificeerd en genetische isolatie kan groeien (Frankham et al., 2002; Bowler et al., 2005). Negatieve effecten van habitatfragmentatie zijn vanzelfsprekend van toepassing op zowel de kleine marterachtigen als die van hun prooidiersoorten. Omwille van een hogere trofische positie kunnen deze echter ernstiger zijn voor de wezel en de hermelijn (Van Nouhuys, 2005). Daartoe geldt
15 verdroging van het landschap potentieel als een extra verzwarende factor voor de verdere degradatie en/of fragmentatie van vochtige habitattypes in het geval van de hermelijn.
Knaagdieren, voornamelijk de bruine rat (Rattus norvegicus), worden vervolgd om economisch verlies te beperken en de volksgezondheid te beschermen in Vlaanderen. De belangrijkste middelen om deze populaties te controleren zijn rodenticiden op basis van anticoagulantia (AR) (Baert et al., 2015). Een nadeel is dat ook andere knaagdieren hieraan worden blootgesteld (Elliott et al., 2014). Daarenboven zijn nieuwe generaties rodenticiden meer toxisch als reactie op een zich ontwikkelde resistentie bij de bruine rat. Niet alleen heeft dit een invloed op de knaagdierpopulatie, maar is de kans op secundaire intoxicatie op hogere trofische niveaus sterk aanwezig (Baert et al., 2015). Eén van de risicofactoren met betrekking tot secundaire intoxicatie is de specificiteit van het dieet in relatie tot knaagdieren (López-Perea & Mateo, 2018). Uit een experimentele setting blijkt dat een concentratie van 0.2µg/g rodenticiden reeds interfereert met de overleving van de hermelijn (Grolleau et al. 1989). Deze concentratie van AR’s werden al vastgesteld bij de hermelijn na een toxicologisch leveronderzoek door Elmeros et al. (2011).
Met de voortschrijdende regularisatie van de jacht en toepassing van beschermingsmaatregelen zijn middelgrote carnivoren en dag- en nachtroofvogels, waaronder de buizerd (Buteo buteo), terug meer verspreid in Vlaanderen (Van Den Berge, 1995; Herremans, 2007; Van Den Berge, 2017). Daarnaast is ook de huiskat (Felis silvestris catus) zeer prominent vertegenwoordigd in het landschap (Van Den Berge et al., 2011). Zoals reeds vermeld, is het niet gekend in welke mate de aanwezigheid van grotere predatoren een impact heeft op de populatiedynamiek van de kleine marterachtigen. Omtrent een eventueel bijkomend effect ten gevolge van een hogere aanwezigheid van deze predatoren blijft het dan ook gissen.
Een andere factor die vaak wordt vermeld als mogelijke bedreiging is een parasitaire besmetting, meer bepaald met Skrjabingylus nasicola.
Skrjabingylus nasicola
Skrjabingylus is een genus van parasitaire rondwormen die zich met behulp van een tussengastheer nestelt
in de frontale sinusholten van de marterachtigen (Hansson, 1968; Anderson, 2000; Koubek et al., 2004; Kierdorf et al., 2006). Het geslacht telt zes soorten waarvan Skrjabingylus nasicola tot op heden de enige soort van deze orde is die wordt vastgesteld als parasiet bij de wezel en de hermelijn (Koubek et al., 2004; Heddergott et al., 2016). Het ziektebeeld manifesteert zich gradueel na vermoedelijk een opeenvolging van besmettingen en uit zich door een vervorming en perforaties van het botweefsel ter hoogte van de supraorbitale regio (McDonald & Lariviere, 2001; King & Powell, 2006). Larven van de nematode worden in de omgeving verspreid via het defecatieproces van de eindgastheer en worden vervolgens opgenomen door terrestrische mollusken. In deze obligate tussengastheer ontwikkelen de larven zich gedurende 12 tot 18 dagen tot een volgend stadium (Dubnitskii, 1956). Rechtstreekse infectie door het consumeren van (naakt)slakken werd in een experimentele setting aangetoond (Dubnitskii, 1956), maar gebaseerd op de hoge prevalentiecijfers dient infectie logischerwijs via paratenische tussengastheren te verlopen. Terrestrische mollusken worden immers niet of slechts zelden aangetroffen in het dieet van de kleine marterachtigen (King & Powell, 2006). De aanwezigheid van Skrjabingylus nasicola werd bevestigd in onder meer spitsmuizen, de rosse woelmuis en de gewone bosmuis die na ingestie van een geïnfecteerde mollusk drager werden (Hansson, 1967; Weber & Mermod, 1983). Eenmaal de larven de eindgastheer bereiken, migreren deze doorheen de abdominale wand richting de frontale sinusholten waarna maturatie wordt bereikt en reproductie plaatsvindt (Dubnitskii, 1956) (figuur 5).
16 Infectie door Skrjabingylus nasicola wordt ononderbroken vastgesteld doorheen het verspreidingsgebied van de kleine marterachtigen (Lewis, 1967; Debrot & Mermod, 1981; King & Moody, 1982). De infectiegraad vertoont hierbij een grote mate van ruimtelijke en temporele variatie (King & Powell, 2006). Op basis van beschadigde schedels worden in Europa prevalentiecijfers beschreven gaande van 20 tot 100% in het geval van de wezel (Lavrov, 1944; Vik, 1955; Hansson, 1970; Van Soest et al., 1972; King, 1977). De klimatologische regio speelt hierbij een belangrijke mediërende rol aangezien de larven van
Skrjabingylus nasicola, wanneer aanwezig in de buitenwereld, gevoelig zijn voor uitdroging en koude
temperaturen. Bijgevolg wordt algemeen een hogere mate van infectie vastgesteld indien sprake is van vochtige en milde klimatologische condities (Hansson, 1974). Daarnaast vormt de plaatselijke voedselecologie van de wezel en hermelijn een tweede mediërende factor in het verklaren van de prevalentiegraad. Zo beschreven Debrot & Mermod (1981) een plotse toename in het aantal besmettingen bij de hermelijn nadat de populatie woelratten crashte. Als reactie op een lager prooiaanbod verbreedde het dieet met inclusie van prooisoorten gekenmerkt door een minder vegetarisch dieet zoals bijvoorbeeld de rosse woelmuis (Debrot & Mermod, 1981).
Studies konden tot nu toe geen effect aantonen op de lichaamsconditie van zowel de wezel als de hermelijn (Simpson et al., 2016). Daarnaast konden populatieafnames evenmin met de parasiet in verband worden gebracht (King & Powell, 2006). Het effect op het individueel niveau wordt hierdoor echter niet ontkracht. Een verminderd schedelvolume wordt aangetroffen bij de Amerikaanse nerts (Neovison vison), leidend tot een verhoogde druk op de hersenen (Bowman & Tamlin, 2007). Bovendien worden gedragsveranderingen bij meerdere diersoorten in associatie gebracht met infectie door andere leden van de rondwormfamilie (Prigioni & Boria, 1995; Hughes et al., 2018). In een experimentele setting toonden stinkdieren (Mephitidae) die werden geïnfecteerd met Skrjabingylus chitwoodorum tekenen van lethargie en een gebrek aan coördinatie. Daarnaast werden ook hersenbloedingen en éénmaal een gedeeltelijke verlamming vastgesteld (Lankester & Anderson, 1971; Hughes et al. 2018).
Figuur 5: links: levenscyclus van Skrjabingylus nasicola met: a) verspreiding van larven via het defecatieproces, b) opname door
een obligate tussengastheer, c) Rechtstreekse besmetting. d) opname door een paratenische tussengastheer en e) besmetting via een paratenische tussengastheer (King & Powell, 2006). Rechts: botperforatie ter hoogte van de supraorbitale regio ten gevolge van S. nasicola (Verschuren, 2005).
17
Monitoring en citizen science
De mogelijkheid om veranderingen in populatiegroottes te detecteren is van fundamenteel belang om risicosoorten te identificeren. Daarnaast biedt het ook inzicht in de ecologie van soorten zodanig efficiënte beschermingsmaatregelen kunnen worden geïdentificeerd en na implementatie ook kunnen worden geëvalueerd (Kleiman et al., 2000; Tear et al., 1995).
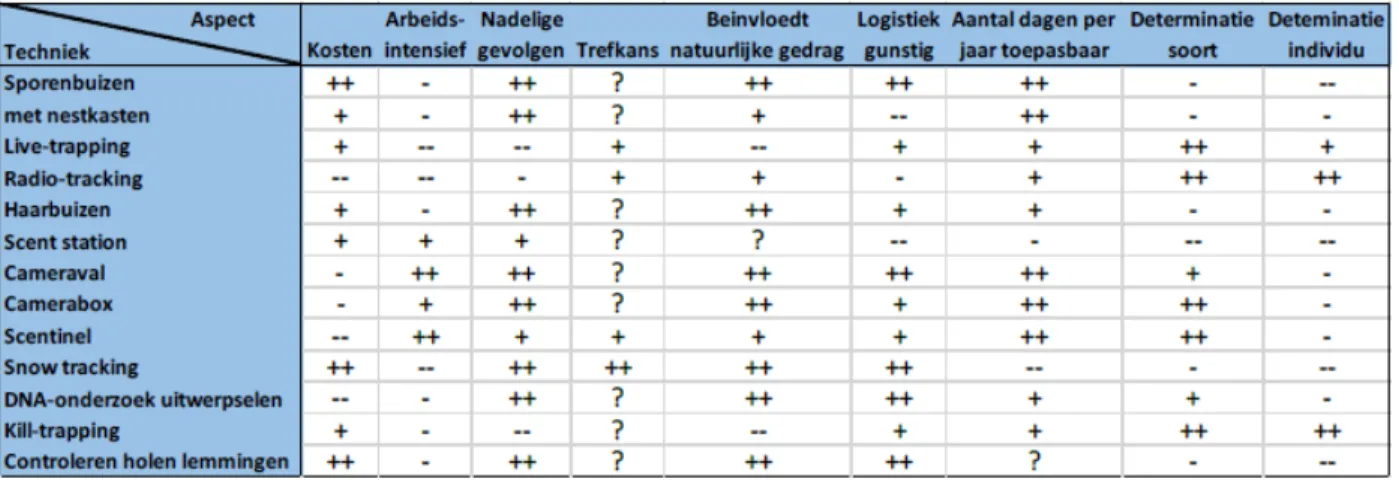
Omwille van de soortkarakteristieke kenmerken van de wezel en de hermelijn is het monitoren van deze soorten echter een grote uitdaging (Van Den Berge & De Pauw, 2003). De meest gebruikte methode in klassiek onderzoek omtrent onder meer habitatvoorkeur, namelijk sporenwaarnemingen, zijn in een Vlaamse context amper of niet toepasbaar door de afwezigheid van een jaarlijks sneeuwlandschap. Daarnaast is de meest succesvolle methode in het verleden, jachtvangsten, niet meer toegelaten. In een literatuuronderzoek uitgevoerd door Van Tongeren (2017) werden mogelijke technieken besproken aan de hand van de toepasbaarheid (figuur 6). Voor een beschrijving van al deze technieken wordt dan ook doorverwezen naar desbetreffend artikel. De auteur concludeert dat voornamelijk het gebruik van sporenbuizen en cameraboxen de meest geschikte methoden zijn voor de Nederlandse, en aldus de Vlaamse situatie. Eén van de vermelde nadelen van sporenbuizen is daarentegen de overlap van pootgrootte bij mannetjeswezels en vrouwtjeshermelijnen (Vuurde & Van der Grift, 2005). Daarnaast hebben deze methoden slechts een reikwijdte met betrekking tot het detecteren van veranderingen in populatiegroottes op een landschapsschaal, tenzij een enorme hoeveelheid aan middelen beschikbaar zou zijn.
Figuur 6: overzicht van het literatuuronderzoek uitgevoerd door Van Tongeren (2017) met betrekking tot monitoringstechnieken.
Citizen science betekent voor het ecologisch onderzoek echter potentieel een stap vooruit in het verzamelen van data op landschapsschaal (Schmeller et al., 2009). Www.waarnemingen.be is een online natuurplatform dat vrijwilligers de kans geeft losse waarnemingen in België te delen. ‘Losse waarnemingen’ of ‘opportunistische waarnemingen’ worden gedefinieerd als waarnemingen die niet volgens een gestandaardiseerd telprotocol hebben plaatsgevonden en bijgevolg enkel informatie bevatten omtrent aanwezigheid (Van Strien et al., 2013). Het gebruik van deze data in het bepalen van een eventuele populatietrend ligt dan ook niet voor de hand omwille van verschillende factoren. In de eerste plaats is in vergelijking met een gestandaardiseerd telprotocol de zoekinspanning in ruimte en tijd niet gekend (Van Strien et al., 2013). Daarnaast verschillen waarnemers ten opzichte van elkaar met betrekking tot ervaring en interesses leidend tot een bijkomende bias in rapportering en detectie (Mackenzie et al., 2017). Bovendien kunnen deze ook voor de waarnemer zelf veranderen na verloop van tijd. Deze
18 beperkingen kunnen aan de basis liggen van incorrecte trends en bijgevolg, de waarde van citizen science-data in het kader van monitoring indammen.
Ingevoerde waarnemingen in het dataportaal www.waarnemingen.be zijn daarentegen gekoppeld aan een profiel. Herremans (2010) gebruikt het activiteitspatroon van deze profielen om de geleverde zoekinspanning te benaderen. Hiertoe wordt het jaarlijks aantal daghokbezoeken van waarnemers gesommeerd. Eén daghokbezoek staat voor één waarnemer die op één dag minstens één waarneming meldt uit één UTM1-hok (1x1km). Daarnaast kan op basis van een selectie van waarnemingsprofielen verder gecorrigeerd worden voor een meer precieze benadering van de zoekinspanning en anderzijds rapporteringsbias limiteren (Herremans & Gielen, 2009).
19
Doelstellingen
Onderhavig thesisonderzoek heeft in de eerste plaats tot doel de resultatenpool, verzameld door Verschuren (2005) verder uit te breiden. Dit gebeurt aan de hand van een nieuwe reeks autopsies verricht op de door het Marternetwerk ingezamelde dode exemplaren sinds 2005. Deze kaderen in het verbreden van de soortspecifieke kennis van de wezel en de hermelijn in Vlaanderen zodanig beide soorten beter gesitueerd kunnen worden ten opzichte van de internationale, wetenschappelijke literatuur. De kennisvergaring moet toelaten mogelijke beschermingsmaatregelen te identificeren indien de conservatiestatus van de wezel of de hermelijn verder achteruit zou gaan in de toekomst. Daarnaast wordt ook aandacht geschonken aan lichaamsgrootte (deels in relatie tot monitoring) en variatie in vachtkarakteristieken. Tenslotte wordt in het huidige onderzoek de kans gegrepen om de mogelijkheden van citizen science met betrekking tot monitoring te exploreren via het online natuurplatform www.waarnemingen.be dat een samenwerking is tussen Natuurpunt, Natagora en Stichting Natuurinformatie (figuur 7).
Figuur 7: leidraad van onderhavig thesisonderzoek.
Autopsiegedeelte
Doodsoorzaak en conditieparameters
Hoewel het Marternetwerk focust op verkeersslachtoffers, beperkt de inzameling van karkassen zich hier niet toe. Andere doodsoorzaken worden vastgesteld/bevestigd en bieden een verdere inkijk in mogelijke bedreigingsfactoren in associatie met het voortbestaan van de kleine marterachtigen in Vlaanderen. Daarnaast wordt ook de algemene conditie van het dier getoetst en in relatie gezet met de doodsoorzaak.
Variatie in vachtkarakteristieken
In navolging van Verschuren (2005) wordt de variatie in vachtkarakteristieken bij beide soorten in Vlaanderen verder gesitueerd.
20
Leeftijdscohorten
Karkassen van beide soorten worden aan een leeftijdscohort toegewezen. Dit onderdeel beperkt zich niet tot de huidige reeks ingezamelde exemplaren, maar neemt ook deze van het vorige onderzoek in rekening. Het benaderen van de leeftijd heeft niet tot doel een populatieverdeling te maken, maar biedt onder meer een inzicht in de temporele spreiding van juvenielen en zodoende, de breedte van de reproductieperiode. Daarenboven worden deze resultaten ook voor andere modules gebruikt (zie verder).
Lichaamsgrootte en seksueel dimorfisme
De lichaamsgrootte en het seksueel dimorfisme van beide kleine marterachtigen varieert sterk doorheen het verspreidingsgebied (Erlinge, 1987; King & Powell, 2006, Yom-Tov et al., 2010). In dit onderzoek worden het lichaamsgewicht en verschillende afmetingen van het lichaam en de schedel bepaald om deze voor de Vlaamse populatie te kaderen. Hiertoe worden karkassen geselecteerd aan de hand van een leeftijdscohort zodanig dat niet-volgroeide dieren worden uitgesloten. Na een selectie van de resultaten van Verschuren (2005), worden deze aan de huidige dataset toegevoegd.
Reproductie
Naast de resultaten bekomen door het toewijzen van leeftijdscohorten, wordt het voortplantingsseizoen van beide soorten verder gesitueerd aan de hand van de reproductiestatus van mannetjes en vrouwtjes, rekening houdend met het toegewezen leeftijdscohort. Voor de mannetjes worden de resultaten van Verschuren (2005) toegevoegd na het bepalen van de leeftijdscategorie.
S. nasicola
De aanwezigheid van de parasitaire rondworm S. nasicola wordt aan de hand van twee methoden in kaart gebracht. De eerste focust op het aantal actuele besmettingen en de tweede heeft tot doel het aantal schadegevallen te kwantificeren bij de populatie behorend tot leeftijdscohort “1” van beide soorten. Een leeftijdsopdeling werd gemaakt aangezien schade progressief optreedt (Dubay et al., 2014) en een grote aanwezigheid van jonge exemplaren een sterk vertekend beeld zou kunnen geven.
Dieet
Voedselbeschikbaarheid wordt beschouwd als de belangrijkste determinant van aan-of afwezigheid van beide kleine marterachtigen (King & Powell, 2006). Omtrent het dieet van de wezel en de hermelijn in Vlaanderen is kennis echter gelimiteerd tot de dataset van Verschuren (2005). Om een ruimer beeld te krijgen wordt de maaginhoud van de ingezamelde exemplaren gedetermineerd en worden deze resultaten gebundeld met die van de vorige dataset. Vervolgens wordt het dieet zowel per soort als tussen beide soorten geanalyseerd.
Pootafmetingen wezel en hermelijn
Monitoring gebruikmakend van sporenbuizen zou gehinderd worden door overlap in pootafdrukken tussen wezelmannetjes en hermelijnvrouwtjes (Vuurde & van der Grift, 2005). Om een antwoord te bieden op deze problematiek wordt in dit onderdeel een aanzet gegeven tot het in kaart brengen van de pootdimensies van beide soorten.
Toekomstig onderzoek
Om mogelijke genetische effecten van habitatfragmentatie na te gaan en de eventuele aanwezigheid van secundaire intoxicatie te controleren werden van elk exemplaar DNA- en weefselstalen (lever, nieren en mesenteriaal vet) genomen en gearchiveerd in het kader van toekomstig onderzoek.
21
Citizen science
Citizen science groeit reeds enkele jaren in Vlaanderen en betekent een mogelijk waardevol alternatief om de populatiegrootte van beide kleine marterachtigen te monitoren. Na een initiële exploratie van het online natuurplatform www.waarnemingen.be wordt in een volgende stap op basis van ingevoerde zichtwaarnemingen meerdere populatietrends berekend, waarbij wordt getracht te corrigeren voor zoekinspanning in tijd en ruimte aan de hand van de methode beschreven door Herremans (2010). Deze methode wordt verder verfijnd door een selectie uit te voeren op het individuele waarnemingsprofiel zodoende zoekinspanning mogelijk beter wordt benaderd en rapporteringsbias potentieel kan beperkt worden. Om de trends te situeren worden deze vervolgens vergeleken met deze van twee referentiesoorten, namelijk de egel (Erinaceus europaeus) en de vos. Hoewel de egel volgens de IUCN Rode Lijst van de Zoogdieren in Vlaanderen geen daling in verspreiding vertoont (Maes et al., 2014), heerst toch het vermoeden dat de soort recent achteruit zou gaan in analogie met andere West-Europese landen (Huijser & Berger, 2000; Hof et al., 2012; Pettett et al., 2018). Om deze reden wordt voor dit onderzoek een dalende trend voor de egel voorspeld ter referentie. Voor de vos wordt daarentegen een stabiel populatiepatroon verwacht. Tenslotte worden de berekende trends vergeleken op basis van het officiële vosafschot en het aantal broedgevallen van de kerkuil, een specialistische predator op woelmuizen (Lefebvre & Vercayie, 2017).
22
Materiaal en methoden
Autopsiegedeelte:
INBO-protocol
In het kader van het Marternetwerk staan verspreid over Vlaanderen een veertig-vijftigtal diepvriezers ter beschikking om lokaal gemelde dode exemplaren van de wezel en de hermelijn tijdelijk te stockeren. Het inzamelen door vrijwilligers gaat gepaard met een formulier waarop datum en plaats dienen genoteerd te worden met extra ruimte voor de vermoedelijke doodsoorzaak en/of andere relevante informatie. In een volgende stap worden deze gecentraliseerd in het Instituut voor Natuur- en Bosonderzoek te Geraardsbergen en bewaard onder ca. -20°C om uiteindelijk, in dit onderzoek, aan een autopsie onderworpen te worden.
De uitvoering van de autopsies volgt in de eerste plaats een door het INBO gestandaardiseerd protocol en voor een uitvoerige beschrijving hiervan wordt doorverwezen naar Verschuren (2005). Dit protocol werd verder uitgebreid met de haaridentificatie van prooiresten, het opmeten van pootdimensies en het bepalen van het leeftijdscohort.
Algemene situering
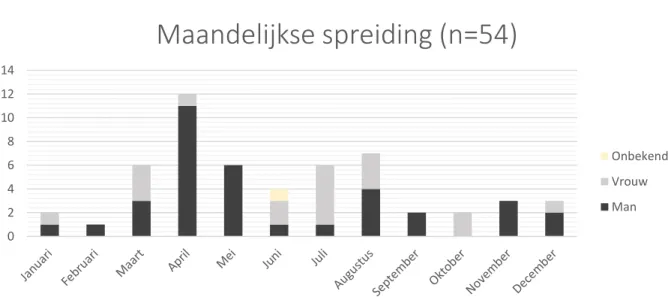
Alle exemplaren geautopseerd tijdens het huidige onderzoek werden in dit onderdeel gesitueerd per maand en per jaar en uitgesplitst per geslacht.
Doodsoorzaken in relatie met conditieparameters
Na een inwendige en uitwendige controle van het kadaver werd de vermelde doodsoorzaak geverifieerd. Hiertoe werd onder meer gefocust op de aanwezigheid van breuken en vachtletsels zoals bijtwonden. Drie conditieparameters werden bepaald:
• Infectie van de longen: kleine stukjes weefsel van de longkwab werden afgeknipt en vervolgens in een glas water gebracht. Het zinken van deze longstukjes wijst op met ontstekingsvocht en etter gevulde alveolen en aldus een longinfectie. Drijvende longstukjes wijzen op het afwezig zijn van een infectie. Indien geen duidelijk resultaat werd geobserveerd, werd de longtest aangeduid als ‘onduidelijk’.
• Niervet: De hoeveelheid vet dat de nieren omgeeft werd visueel beoordeeld volgens één van volgende categorieën: ‘geen tot weinig’, ‘matig tot veel’, ‘volledig omgeven’.
• Onderhuids vet: de hoeveelheid onderhuids vet werd visueel beoordeeld na het losmaken van de huid ter hoogte van de buik- en liesregio. De volgende categorisatie werd gehanteerd: ‘geen tot weinig’, ‘matig tot veel’, ‘volledig omgeven’.
De doodsoorzaken werden voor beide soorten overlopen en uitgesplitst per geslacht. In een volgende stap werden deze geassocieerd met de beschreven conditieparameters.
Variatie in vachtkarakteristieken
Voor de wezel werd per exemplaar gecontroleerd of de demarcatielijn een grillig of recht patroon vertoonde (figuur 8). Daarnaast werd bepaald of de vacht gekenmerkt werd door de aanwezigheid van bruine eilandjes ter hoogte van de witte vacht, aanwezigheid van witte eilandjes ter hoogte van de bruine vacht of afwezigheid van eilandjes (figuur 8). Ook werd de kleur van de voor- en achterpoten beoordeeld, namelijk ‘volledig bruine poten’, ‘volledig witte poten’ of ‘bruine poten met witte tenen’ (figuur 8). Deze
23 resultaten werden uitgesplitst per geslacht. Met betrekking tot de hermelijn werd in de eerste plaats de variatie in vachttypes bepaald voor de periode november-mei aan de hand van de volgende categorisatie: ‘boven bruin, onder wit’, ‘wit-bruin gevlekt’ en ‘wit’ (figuur 9). Hierbij werd geen onderscheid gemaakt tussen een wintervacht of een overgangsvacht met betrekking tot het ‘wit-bruin gevlekt’ vachttype. Ten tweede werd de variatie aangaande de kleur van de ventrale vacht, ‘wit’ of ‘geelwit’ in het geval van het vachttype ‘boven bruin, onder wit’ uitgelicht en vergeleken per geslacht.
Figuur 8: een mannetjeswezel met witte tenen aan de voorpoot, volledig bruine achterpoten, bruine eilandjes ter hoogte van de
hals (naast de typische wangvlekken) en met een grillige demarcatielijn.
24
Leeftijdscohorten
Gebruikmakend van de DSB-methode (Datum, Schedel en Baculum/Penisbeen) (King, 1980 b; King, 1991), werden individuen toegewezen aan een leeftijdscohort (figuur 10). Als eerste stap werden schedels, schedelfragmenten en/of bacula opgedeeld per sterftemaand waarna op basis van meerdere leeftijdsafhankelijke kenmerken een onderscheid werd gemaakt tussen twee leeftijdscategorieën. Voor een korte bespreking van deze kenmerken wordt doorverwezen naar de appendix.
Voor de wezel werden volgende leeftijdscohorten gedefinieerd:
• Leeftijdscohort “0” omvat alle exemplaren geboren in het sterftejaar. • Leeftijdscohort “1” omvat alle exemplaren geboren vóór het sterftejaar. Voor de hermelijn werden volgende leeftijdscohorten gedefinieerd:
• Leeftijdscohort “0” omvat alle exemplaren jonger dan één jaar waarbij 1 april als geboortedatum werd beschouwd.
• Leeftijdscohort “1” omvat alle exemplaren ouder dan één jaar waarbij 1 april als geboortedatum werd beschouwd.
Exemplaren die op basis van de schedel of het baculum niet tot een cohort konden worden toegewezen, werden ondergebracht in de categorie ‘N.T.B.’ (niet te bepalen). De resultaten werden voor beide soorten per geslacht opgesplitst en voor de mannetjes werd baculumgewicht (0.00001 gram) uitgezet in functie van de dag van het jaar en opgesplitst per gedefinieerd cohort.
Voor dit onderdeel werden de schedels en bacula uit het voorgaande onderzoek opnieuw geanalyseerd en toegevoegd aan de huidige dataset.
De gebruikte schedels in deze werden na het verwijderen van het lichaam in met water gevulde glazen gelegd waarna deze ongeveer vier dagen in een warmwaterbad werden geplaatst (70°C). Vervolgens werden deze gespoeld boven een zeef en aan de lucht gedroogd. De gebruikte bacula in dit onderdeel werden na het verwijderen van het lichaam gedurende één tot twee uur in een erlenmeyer op een hete kookplaat gezet, waarna deze aan de lucht werden gedroogd.
25 Figuur 10: DSB-methode in actie voor de wezel tijdens de maand juli: de zes exemplaren links op de foto werden allen toegewezen
tot cohort “0” en de overige twee tot cohort “1”.
Lichaamsgrootte en seksueel dimorfisme
Lichaamsafmetingen en -gewichtenFysiek adulte exemplaren werden geïdentificeerd op basis van het leeftijdscohort. Voor de wezelvrouwtjes werden enkel exemplaren weerhouden behorend tot leeftijdscohort “1”, waardoor een minimumleeftijd van 3-4 maanden werd gegarandeerd indien september als laatste geboortemaand wordt beschouwd. Voor mannetjes werden enkel exemplaren van leeftijdscohort “1” weerhouden exclusief januari en februari zodanig dat een minimum leeftijd van 5-6 maanden was verzekerd indien september als laatste geboortemaand wordt beschouwd.
Bij de hermelijn werden enkel mannetjes behorend tot leeftijdscohort “1” geselecteerd waardoor een minimumleeftijd van één jaar werd verzekerd indien 1 april als gemiddelde geboortedatum wordt beschouwd. Voor de vrouwtjes werden naast exemplaren behorend tot cohort “1” ook exemplaren van cohort “0” en onbepaalde exemplaren weerhouden vanaf oktober. Hierdoor werd een minimumleeftijd van zes maanden gegarandeerd indien 1 april als gemiddelde geboortedatum wordt beschouwd.
Volgende lichaamsafmetingen werden bepaald:
• Kopromplengte: de lengte van de top van de snuit tot de anus gemeten met behulp van een meethoek (1 mm nauwkeurig). Hiertoe werd het karkas plat op de rug gelegd met een natuurlijke strekking en werd de snuit tegen het opwaartse deel van de meethoek geplaatst.
• Staartlengte: de lengte vanaf de anus tot het staarteinde zonder staartpluim gemeten met behulp van een lat (1 mm nauwkeurig).
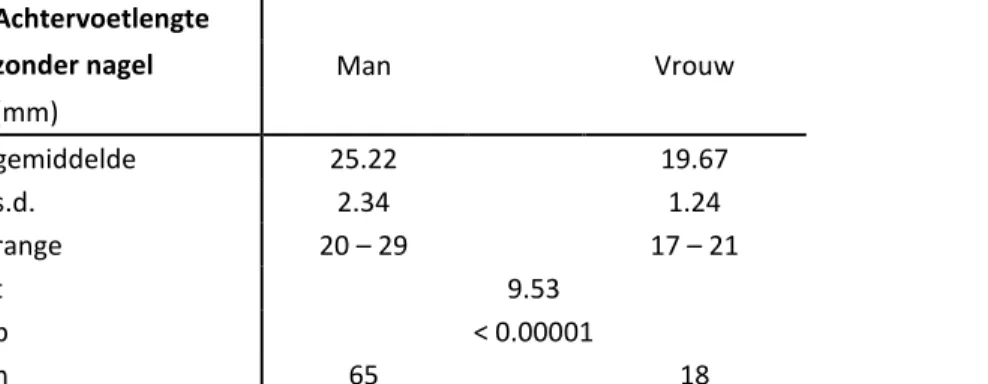
• Achtervoetlengte zonder nagel: de lengte vanaf de hiel tot de top van het hoogste voetkussen gemeten met behulp van een meethoek (1 mm nauwkeurig). Hiertoe werd de hiel tegen de rechte hoek van de meethoek geplaatst.
• Achtervoetlengte met nagel: de lengte vanaf de hiel tot de top van het hoogste voetkussen gemeten met behulp van een meethoek (1 mm nauwkeurig). Hiertoe werd de hiel tegen de rechte hoek van de meethoek geplaatst.
• Oorlengte: de lengte vanaf de groeve onderaan het oor tot het hoogste punt van de oorschelp gemeten met een lat (1 mm nauwkeurig).
26 • Kopomtrek: de omtrek van de kop ter hoogte van de oren gemeten met een meetlint (1mm
nauwkeurig).
• Halsomtrek: de omtrek van de hals gemeten met een meetlint (1 mm nauwkeurig). • Totaal gewicht: het totale gewicht van een intact exemplaar (1 gram nauwkeurig).
• Ontweid gewicht: het gewicht van een intact exemplaar na het verwijderen van de ingewanden, het ademhalingsstelsel, het hart en de reproductieorganen inclusief baculum (1 gram nauwkeurig).
Na de cohortbepaling van de exemplaren geautopseerd door Verschuren (2005), werden de door haar gemeten afmetingen aan dit onderzoek toegevoegd. Voor de gewichtscategorieën werden sterk vermagerde exemplaren met als doodsoorzaak ‘bijvangst’ (in vallen die niet onmiddellijk dodend zijn) niet weerhouden.
Geslachtsverschillen werden statistisch getoetst met de éénzijdige t-test. De resultaten worden voor beide soorten per geslacht weergegeven met vermelding van gemiddelde waarde, range, standaarddeviatie, de t-waarde, significantiewaarde en staalgrootte. Het seksueel dimorfisme werd bepaald aan de hand van de kopromplengte.
Schedelafmetingen
Voor het meten van de schedelafmetingen werden enkel exemplaren weerhouden met de rechte, adulte schedelvorm (zie appendix).
Volgende afmetingen van de schedel werden bepaald met behulp van een digitale schuifpasser (0.01 mm nauwkeurig) (figuur 11):
• Condylobasaallengte (CBL): de afstand tussen de condylus occipitalis of achterhoofdknobbel en het voorste punt van het premaxillare.
• Tandenrijlengte bovenkaak: de afstand tussen de rand van de voorste alveole van de caninus en de achterste rand van de alveole van de laatste molare.
• Tandenrijlengte onderkaak: de afstand tussen de voorste rand van de alveole van de caninus en de achterste rand van de alveole van de laatste molare.
• Onderkaaklengte: de afstand tussen de processus condylus en de voorste rand van de alveole van de incisivi.
• Interorbitaalbreedte: de kleinste afstand tussen de orbita. • Postorbitaalbreedte: de kleinste afstand tussen de parietale.
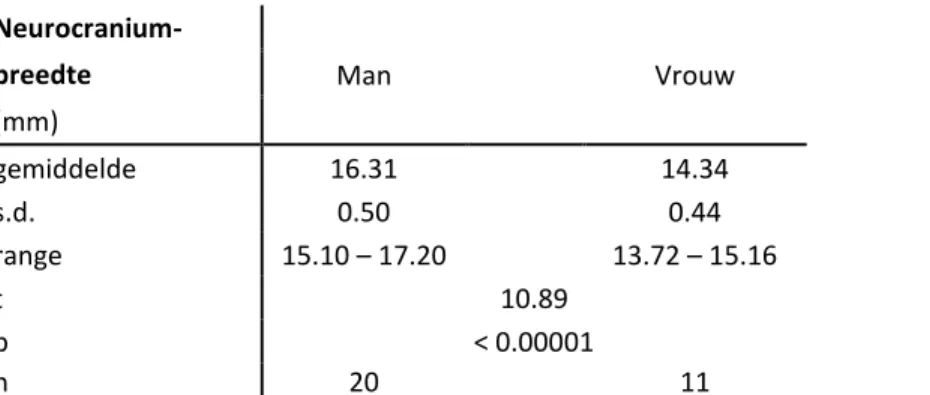
• Neurocraniumbreedte: de kleinste afstand van het neurocranium achter het os squasom.
De gedefinieerde afmetingen werden driemaal herhaald waarna het gemiddelde hiervan werd genomen. De schedels afkomstig van het onderzoek door Verschuren (2005) werden in deze module opnieuw gemeten.
Geslachtsverschillen werden statistisch getoetst aan de hand van de éénzijdige t-test. De resultaten worden voor beide soorten per geslacht weergegeven met vermelding van gemiddelde waarde, range, standaarddeviatie, de t-waarde, significantiewaarde en staalgrootte. Het seksueel dimorfisme werd uitgedrukt aan de hand van de condylobasaallengte.