Biologische schimmelbestrijding
in sla en boon door cyclische
lipopeptide producerende bacteriën
Aantal woorden: 32.663Jarne Depaepe
Stamnummer: 01507935Promotor: Prof. dr. ir. Monica Höfte
Tutor: ir. Rutger Pipeleers
Masterproef voorgelegd voor het behalen van de graad in Master of Science in de bio-ingenieurswetenschappen: landbouwkunde
Deze pagina is niet beschikbaar omdat ze persoonsgegevens bevat.
Universiteitsbibliotheek Gent, 2020.
This page is not available because it contains personal information.
Ghent University, Library, 2020.
v
Voorwoord
Voor een thesisonderwerp was ik vooral op zoek naar een leuk project binnen de gewasbescherming. Een thesis binnen de vakgroep van Fytopathologie sprak me uiteindelijk het meest aan. Na de lessen over Fytopathologie en het gesprek met professor Höfte over de verschillende thesisonderwerpen, was ik verkocht. Het proberen zoeken naar oplossingen voor zeer actuele problemen geeft me als wetenschapper enorm veel voldoening. Mee op zoek gaan naar oplossingen om landbouw te verbeteren is het doel. Biologische bestrijding is hierin een enorm belangrijke factor en in mijn verdere carrière zou ik graag verder gaan in deze sector. Tijdens de practica in mijn eerste master viel het op dat de sfeer in het labo zeer aangenaam was. Na een jaar onderzoek kan ik dit alleen maar bevestigen. Ik maak van dit voorwoord gebruik om enkele mensen te bedanken die deze thesis mee hebben helpen vormen tot wat het geworden is. Ik wil professor Höfte bedanken voor de lessen Fytopathologie, om zo de interesse voor de gewasbescherming op te wekken. Ook voor de algemene begeleiding doorheen het jaar, het bijsturen van de thesis en de kritische blik op de resultaten wil ik u bedanken. Daarnaast wil ik ook mijn rechtstreekse begeleider Rutger bedanken, voor alle tijd die hij investeerde in mij. Ondanks de kleine taalbarrière verliep de communicatie vlekkeloos. Je was altijd meteen te bereiken en legde alle technieken in het labo perfect uit. Wanneer er iets misliep tijdens de proeven bleef je altijd rustig en zochten we samen naar oplossingen. Ook de lange dagen in het labo gingen snel voorbij door toffe gesprekken over Fytopathologie en vele andere zaken. Veel succes met je verdere doctoraat en bedankt om me van goede begeleiding te voorzien.
Ook de andere mensen van de vakgroep Fytopathologie wil ik bedanken. Als iets onduidelijk was kon ik altijd terecht bij jullie met mijn vragen. Ilse en Shirley, heel erg bedankt om mijn laatste grote proef te evalueren en de resultaten door te sturen. Ik vond het heel erg spijtig dat ik dit zelf niet heb kunnen doen, maar corona besliste daar anders over. Door jullie heb ik mijn werk van vijf weken lang niet moeten wegsmijten. Ook wanneer ik last minute vroeg om bepaalde bacteriën uit collectie te halen doorheen het jaar, was dit geen probleem voor Ilse. Aan iedereen bedankt voor de aangename tijd! Verder wil ik zeker en vast mijn ouders specifiek bedanken. Eerst en vooral om me de mogelijkheid te geven om te studeren wat me aansprak. Daarnaast ook voor de onvoorwaardelijke steun tijdens de vijf zware jaren van universitaire studies. Als ik slechtgezind was omdat ik bepaalde leerstof maar niet in mijn hoofd kreeg, lieten jullie me gerust tot ik weer goedgezind was. Daarnaast wil ik ook mijn broer Pieterjan bedanken voor de toffe momenten tijdens de voorbije jaren. Ook aan mijn vriendin Margo, bedankt voor de leuke momenten tussen het thesissen door en de steun doorheen het jaar.
Tot slot zou ik graag mijn vrienden en familie bedanken voor de topmomenten en de interesse over alles waarmee ik bezig ben. Ruben, Elias en Margo, de allerbeste landbouwbuddies die ik me kan voorstellen. Het is fantastisch om dezelfde interesses te delen met jullie en daarnaast nog eens te klikken op zoveel andere vlakken. Ward, Lieven en Arthur, vanaf het eerste jaar bio-ingenieur vormen we een hechte groep en helpen we elkaar waar nodig. Samen de bachelorproef doen met jullie was een hoogtepunt! Milan en Jasper, bedankt voor de legendarische matchen op Club Brugge, de perfecte uitlaatklep. Ook de andere vrienden van mijn dichtste vriendengroep wil ik in het bijzonder bedanken voor de dagelijkse momenten van ontspanning en plezier.
Jarne Depaepe Gent, 2020
vii
Preambule: Covid-19
Begin 2020 brak wereldwijd een pandemie uit door de verspreiding van het coronavirus. Verschillende landen namen maatregelen om de verspreiding tegen te gaan. Ook de Belgische overheid besliste om de mensen zoveel mogelijk thuis te houden en contacten te vermijden. Op 16 maart werd ook de toegang tot de gebouwen van de UGent geweigerd voor alle studenten. Dit verbod had een impact op de thesis aangezien de labo’s niet meer beschikbaar waren om verdere proeven uit te voeren. De proeven die uitgevoerd zijn tussen 1 augustus 2019 en 16 maart 2020 konden doorgaan zoals gepland. Twee proeven die in een eindfase zaten konden geëvalueerd worden door medewerkers uit de vakgroep die wel nog toegang hadden tot het labo. Het gaat concreet over het scoren van de proef op boon na infectie met Rhizoctonia solani en Botrytis cinerea (sectie 3.2.3) en de 2de herhaling van de
biocontrole proef op sla na infectie met Fusarium oxysporum (sectie 3.3.2). Het tijdstip waarop gescoord werd was niet ideaal door de gegeven omstandigheden. Twee andere proeven die nog gepland stonden na 16 maart konden niet meer doorgaan. Hier gaat het om een proef op directe antagonisme bij boon die uitgebreid zou worden na een eerste inleidende proef (sectie 3.2.2). De andere proef kaderde in het onderzoek naar de impact van verschillende bodems op de infectiecapaciteit van Fusarium oxysporum in sla (sectie 3.4.1).
De thesis werd afgewerkt met de beschikbare data uit de verschillende uitgevoerde proeven. Er was geen heroriëntering nodig van de thesis omdat het reeds voorhanden materiaal voldoende was om een volwaardig werk af te leveren.
Deze preambule werd in overleg tussen de student en de promotor opgesteld en door beiden goedgekeurd.
ix
Lijst met afkortingen
CLP Cyclische lipopeptide
Fol Fusarium oxysporum f. sp. lactucae
GBS Genotype-by-sequencing
IAA Indool-3-azijnzuur
IPM Integrated pest management
ISR Induced Systemic Resistance
MAMP Microbe associated molecular patterns PAMP Pathogen associated molecular patterns
PDA Potato dextrose agar
PDB Potato dextrose broth
rpm Revolutions per minute
SAR Systemic acquired resistance WLIP White line-inducing principle
xi
Samenvatting
In de landbouw wordt meer en meer op zoek gegaan naar alternatieve methoden om gewassen te beschermen. Chemische middelen kunnen de gezondheid van de mens en het milieu aantasten, waardoor hun gebruik moet worden ingeperkt. In deze thesis wordt het gebruik van biocontrole bacteriën bestudeerd als alternatieve gewasbeschermingstechniek. Tal van wortel geassocieerde
Pseudomonas en Bacillus bacteriën produceren cyclische lipopeptiden (CLP's) als secundaire
metabolieten. Van deze CLP's is bekend dat ze antifungale activiteit vertonen en in staat zijn om resistentie te induceren in planten. De biocontrole capaciteit van verschillende CLP-producerende bacteriën werd bestudeerd. Deze bacteriën werden onder andere geëvalueerd voor de biocontrole van Fusarium oxysporum in sla en Rhizoctonia solani & Botrytis cinerea in boon. De exacte functie van de CLP's in dit biocontrole proces werd onderzocht via mutanten die niet langer in staat waren om specifieke CLP's te produceren.
In de bonenteelt zijn heel wat pathogenen gekend die de planten kunnen infecteren en zo de oogst drastisch verlagen. In deze thesis werd gebruik gemaakt van Rhizoctonia solani en Botrytis cinerea. Er werd getest op wortelrot veroorzaakt door Rhizoctonia solani AG4 (directe antagonisme) en op ‘web blight disease’ veroorzaakt door de bladpathogeen Rhizoctonia solani AG2-2 (geïnduceerde systemische resistentie - ISR). Bacillus amyloliquefaciens GA1 vertoonde zowel in vitro als in planta potentieel om via directe antagonisme wortelrot te reduceren. Hier werden geen duidelijke invloeden van CLP’s waargenomen. De infectie van het blad met Rhizoctonia solani AG2-2 werd erger na behandeling met B. amyloliquefaciens GA1, wat erop wijst dat deze bacterie geen resistentie kan induceren bij boon tegen deze pathogeen. Pseudomonas putida RW10S2 (WLIP-producent) reduceerde de B. cinerea laesie en stimuleerde de vorming van wortelnodules. Bij de mutant die het CLP WLIP niet kon aanmaken verdwenen zowel de positieve eigenschappen ten opzichte van B. cinerea als de vorming van extra wortelnodules.
De teelt van sla komt de laatste jaren meer en meer in de problemen door de Fusarium verwelkingsziekte. Vooral sinds 2016, toen een nieuwe fysio ontdekt werd van Fusarium oxysporum f.sp. lactucae, hebben telers te maken met enorme verliezen. Chemische middelen ter bestrijding van de pathogeen zijn op dit moment niet beschikbaar. Van de verschillende Pseudomonas en Bacillus stammen die getest zijn, vertoonde Bacillus amyloliquefaciens GA1 zowel in vitro als in planta het meeste potentieel om als biocontrole organisme gebruikt te worden. Vooral de CLP-families van de iturins en de fengycins bleken hiervoor verantwoordelijk. De meeste Pseudomonas species hadden zowel in vitro als in planta geen effect op de groei van Fusarium oxysporum f.sp. lactucae. Enkel P.
koreensis Cow5 en P. putida Cor55, respectievelijk CLP-producenten van cocoyamide A en putisolvin,
vertoonden in vitro een klein effect. In planta verdween dit effect volledig.
Tot slot werd voor enkele zeer nauw verwante soorten van Fusarium oxysporum f.sp. lactucae fysio 4 getest of ze sla konden infecteren. In een eerdere studie werd via multilocus sequenties (tef1, cmdA en rpb2) gevonden dat F. oxysporum f. sp. rhois CBS 220.49, F. oxysporum f. sp. tulipae CBS 242.59, F.
nirenbergiae CBS 130303 en F. curvatum CBS 247.61 zeer gelijkaardig tot niet te onderscheiden waren
van F. oxysporum f. sp. lactucae fysio 4. Uit onze resultaten bleek dat geen van de geteste stammen in staat was om de slaplanten te laten verwelken. Alle stammen werden wel latent teruggevonden in de wortels.
xiii
Abstract
In agriculture, alternative methods to protect our crops are increasingly being investigated. Chemical products can affect human health and the environment, so their use must be limited. An interesting alternative is the use of plant beneficial bacteria to act as biological control agents. Numerous root associated Pseudomonas and Bacillus bacteria produce cyclic lipopeptides (CLPs) as secondary metabolites. These CLPs are known to have antifungal activity. Some of these compounds can also induce resistance in several plants. The capacity as biocontrol agents of several CLP-producing bacteria was studied. These bacteria were evaluated for biocontrol of Fusarium oxysporum in lettuce and
Rhizoctonia solani & Botrytis cinerea in bean. The function of CLPs in this biocontrol process was
investigated by using mutants that were no longer able to produce specific CLPs.
Many pathogens can infect beans and drastically reduce yield. In this thesis root rot caused by
Rhizoctonia solani AG4 (Direct antagonism), and web blight disease caused by foliar pathogen Rhizoctonia solani AG2-2 (Induced systemic resistance) were tested. Bacillus amyloliquefaciens GA1
showed potential to reduce root rot both in vitro and in planta. However, no clear influences of specific CLP families were observed. The infection of Rhizoctonia solani AG2-2 on bean leaves got worse after treatment with B. amyloliquefaciens GA1, which indicates that this bacterium cannot induce immunity in bean. It was also found that Pseudomonas putida RW10S2 (WLIP-producer) could induce a significantly smaller lesion of Botrytis cinerea after colonization of the roots. A higher number of root nodules was found and the leaves turned dark green. In case of the WLIP-mutant of RW10S2, both positive properties against B. cinerea and the formation of additional root nodules disappeared Recently, the cultivation of lettuce in Flanders has become difficult due to Fusarium wilting disease. Especially since 2016 when a new race of Fusarium oxysporum f.sp. lactucae was discovered, growers are facing huge problems. Chemicals to control the pathogen are not available. Of the various
Pseudomonas and Bacillus strains tested, Bacillus amyloliquefaciens GA1 showed the most potential
to be used as a biocontrol organism both in vitro and in planta. Especially the CLP families of the iturins and fengycins proved to be responsible for this activity. Most Pseudomonas species had no effect on the growth of Fusarium oxysporum f.sp. lactucae both in vitro and in planta. Only P. koreensis Cow5 and P. putida Cor55, CLP-producers of cocoyamide A and putisolvin respectively, showed a small effect
in vitro. In planta, this inhibition disappeared completely.
Finally, some very closely related species of Fusarium oxysporum f.sp. lactucae race 4 were tested whether they could infect lettuce. In an earlier study, multilocus sequences (tef1, cmdA and rpb2) showed that F. oxysporum f. sp. rhois CBS 220.49, F. oxysporum f. sp. tulipae CBS 242.59, F.
nirenbergiae CBS 130303 and F. curvatum CBS 247.61 were very similar or even indistinguishable from F. oxysporum f. sp. lactucae race 4. None of these strains was able to show symptoms on lettuce plants
xv
Inhoudstabel
Voorwoord ... v
Preambule Covid-19 ... vii
Lijst met afkortingen ... ix
Samenvatting ... xi
Abstract ... xiii
1. Inleiding en onderzoeksvragen ... 1
2. Literatuurstudie ... 3
2.1 Sla - Fusarium oxysporum f. sp. lactucae ... 3
2.1.1 Inleiding slateelt ... 3
2.1.2 Bedreiging door Fusarium oxysporum ... 4
2.1.3 Bestaande bestrijdingstechnieken tegen Fusarium oxysporum ... 6
2.2 Boon - Rhizoctonia solani & Botrytis cinerea ... 7
2.2.1 Inleiding bonenteelt ... 7
2.2.2 Bedreiging door Rhizoctonia solani ... 8
2.2.3 Bestaande bestrijdingstechnieken tegen Rhizoctonia solani ... 9
2.2.4 Bedreiging door Botrytis cinerea ... 10
2.2.5 Bestaande bestrijdingstechnieken tegen Botrytis cinerea ... 12
2.3 CLP producerende bacteriën als biocontrole oplossing ... 13
2.3.1 Verschuiving van chemische naar biologische bestrijding ... 13
2.3.2 Cyclische lipopeptiden ... 14
2.3.3 Pseudomonas biocontrole organismen ... 16
2.3.4 Bacillus biocontrole organismen ... 22
3. Materiaal en methoden ... 25
3.1 Overzicht bacteriën en opkweek... 25
3.1.1 Gebruikte bacteriën ... 25
3.1.2 Opkweek bacteriën ... 26
3.2 Pathosystemen Boon - Rhizoctonia solani & Botrytis cinerea ... 26
3.2.1 Direct in vitro antagonisme tegen Rhizoctonia solani ... 26
3.2.2 Direct in planta antagonisme tegen Rhizoctonia solani AG4 ... 27
xvi
3.3 Pathosysteem Sla - Fusarium oxysporum f. sp. lactucae ... 31
3.3.1 Direct in vitro antagonisme tegen Fusarium oxysporum ... 31
3.3.2 In planta biocontrole tegen Fusarium oxysporum ... 32
3.4 Invloedsfactoren bij de Fusarium oxysporum infectie van sla ... 33
3.4.1 Infectiecapaciteit van verwante Fusarium spp. ... 33
3.5 Statistische analyse ... 34
4. Resultaten ... 35
4.1 Pathosystemen Boon - Rhizoctonia solani & Botrytis cinerea ... 35
4.1.1 Direct in vitro antagonisme tegen Rhizoctonia solani ... 35
4.1.2 Direct in planta antagonisme tegen Rhizoctonia solani AG4 ... 38
4.1.3 Geïnduceerde systemische resistentie (ISR) door Pseudomonas en Bacillus ... 39
4.2 Pathosysteem Sla - Fusarium oxysporum f. sp. lactucae ... 45
4.2.1 Direct in vitro antagonisme tegen Fusarium oxysporum ... 45
4.2.2 In planta biocontrole tegen Fusarium oxysporum ... 48
4.3 Invloedsfactoren bij de Fusarium oxysporum infectie van sla ... 50
4.3.1 Infectiecapaciteit van verwante Fusarium spp. ... 50
5. Discussie ... 53
5.1 Pathosystemen Boon - Rhizoctonia solani & Botrytis cinerea ... 53
5.1.1 Direct in vitro antagonisme tegen Rhizoctonia solani ... 53
5.1.2 Direct in planta antagonisme tegen Rhizoctonia solani AG4 ... 55
5.1.3 Geïnduceerde systemische resistentie (ISR) door Pseudomonas en Bacillus ... 56
5.2 Pathosysteem Sla - Fusarium oxysporum f. sp. lactucae ... 59
5.2.1 Direct in vitro antagonisme tegen Fusarium oxysporum ... 59
5.2.2 In planta biocontrole tegen Fusarium oxysporum ... 60
5.3 Invloedsfactoren bij de Fusarium oxysporum infectie van sla ... 61
5.3.1 Infectiecapaciteit van verwante Fusarium spp. ... 61
6. Algemene conclusie en verder onderzoek ... 62
7. Referenties ... 64
1
1. Inleiding en onderzoeksvragen
Sla en boon zijn economisch belangrijke gewassen in Vlaanderen. De teelten krijgen steeds meer te maken met pathogenen waardoor het behalen van een kwalitatieve oogst bemoeilijkt wordt. Het gebruik van bodemontsmetters als dazomet wordt steeds strenger gereguleerd tot zelfs volledig verboden. Het gebruik van chemische pesticiden is niet altijd succesvol en kadert bovendien niet in een duurzaam landbouwsysteem. Daarnaast moeten telers rekening houden met een maximum aantal residuen die aanwezig mogen zijn in de gewassen, ook op vraag van retailers en consumenten. De Europese Unie zet ook volop in op een duurzaam landbouwmodel met Integrated Pest Management (IPM), waar chemische middelen slechts in het laatste geval gebruikt mogen worden. Het is dan ook de bedoeling om alternatieve en duurzame methodes te zoeken om de pathogenen terug te dringen. In deze thesis worden cyclische lipopeptide producerende bacteriën als Pseudomonas en Bacillus bestudeerd als veelbelovende biocontrole organismen.
Eerst en vooral werd een literatuurstudie uitgevoerd om de algemene onderwerpen toe te lichten. Hierbij werd alle informatie over de onderzochte pathosystemen, de potentiële biocontrole organismen en de cyclische lipopeptiden gebundeld.
Twee van de drie onderzochte pathosystemen waren Rhizoctonia solani-boon en Botrytis cinerea-boon. R. solani zorgt onder andere voor wortelrot (infectie via wortels) en ‘web blight disease’ (infectie via het blad). Om wortelrot te evalueren werd gebruik gemaakt van de stam R. solani AG 4-HGI CuLT-Rs36 (Nerey et al., 2010), geïsoleerd in Cuba. Deze stam staat bekend als een agressieve veroorzaker van wortelrot. Voor ‘web blight disease’ werd R. solani AG2-2 CuHav-Rs18 (Nerey et al., 2010) gebruikt, eveneens geïsoleerd in Cuba. Er werd onderzocht of via Pseudomonas en Bacillus soorten, die wortels kunnen koloniseren, aan biologische controle kan gedaan worden. Hierin spelen de cyclische lipopeptiden (CLP’s) die ze produceren wellicht een grote rol. Via zowel in vitro als in planta proeven met mutanten werd nagegaan welke CLP’s een invloed spelen bij een eventuele inhibitie. Zowel directe antagonisme ten opzichte van de pathogeen (R. solani AG4) als het induceren van een verhoogde resistentie in de plant (R. solani AG2-2) werd onderzocht. Daarnaast werd bij boon ook biocontrole tegen de pathogeen B. cinerea bestudeerd. Deze studie focuste vooral op het induceren van verhoogde resistentie in de planten. Voor de infectie met Botrytis werd gebruik gemaakt van de stam B. cinerea R16 (Faretra & Pollastro, 1993).
Het laatste onderzochte pathosysteem was Fusarium oxysporum-sla, wat voor grote problemen zorgt in de Vlaamse en Nederlandse slateelt. Van deze verwelkingsziekte werd fysio 4 pas recent ontdekt waardoor weinig bestaande bestrijdingstechnieken gekend zijn. Het gebruikte isolaat voor alle proeven was Fus1.02 (Claerbout, 2020). Via Pseudomonas en Bacillus bacteriën werd in deze thesis onderzocht of de bacteriën via wortelkolonisaties in staat zijn om Fusarium oxysporum f. sp. lactucae te onderdrukken. Hiervoor werden zowel testen in vitro als in planta uitgevoerd. Via mutanten kon worden onderzocht welke CLP’s verantwoordelijk waren voor een eventuele inhibitie via directe antagonisme.
2
Tot slot werden factoren onderzocht die mogelijk een impact hebben op de infectie van sla door
Fusarium oxysporum. In een eerste deel werd de rol van de pathogeen in het infectieproces
bestudeerd. Claerbout (2020) vond via multilocus sequenties (tef1, cmdA en rpb2) dat F. oxysporum f. sp. rhois CBS 220.49, F. oxysporum f. sp. tulipae CBS 242.59, F. nirenbergiae CBS 130303 en F. curvatum CBS 247.61 zeer gelijkaardig tot niet te onderscheiden waren van F. oxysporum f. sp. lactucae fysio 1 en 4. In deze thesis werd onderzocht of deze genetisch nauw verwante stammen ook in staat waren om sla te infecteren. In een tweede deel zou gekeken worden of de infectie van Fol beïnvloed wordt door het bodemtype. Om dit na te gaan werd sla opgekweekt in verschillende gronden waaraan eenzelfde dosis Fol werd toegevoegd. Na drie weken werden de verwelkingssymptomen geëvalueerd als maat voor de Fol infectie. Daarnaast was het de bedoeling om wortel koloniserende bacteriën uit deze gronden te isoleren. De proeven rond de verschillende gronden heeft niet kunnen plaatsvinden wegens de maatregelen die genomen werden in het kader van de coronacrisis (zie preambule Covid-19).
Voor deze thesis worden volgende onderzoeksvragen geformuleerd:
Onderzoeksvraag 1: Welke Pseudomonas en Bacillus bacteriën kunnen gezien worden als potentiële biocontrole organismen voor de bestrijding van Rhizoctonia solani en Botrytis cinerea bij boon? Welke CLP’s spelen een rol bij het onderdrukken van deze schimmelpathogenen?
Onderzoeksvraag 2: Welke Pseudomonas en Bacillus bacteriën vertonen potentieel als biocontrole organismen voor de bestrijding van Fusariumverwelking in sla? Welke CLP’s hebben een invloed op de inhibitie van de schimmelpathogeen?
Onderzoeksvraag 3: Kunnen zeer nauw verwante stammen van Fusarium oxysporum f. sp. lactucae fysio 4 ook verwelking veroorzaken in sla?
3
2. Literatuurstudie
2.1 Sla - Fusarium oxysporum f. sp. lactucae
2.1.1 Inleiding slateelt
Sla of Lactuca sativa L. behoort tot de composietenfamilie of Asteraceae, ook wel de samengesteldbloemigen genoemd. De productiecijfers wereldwijd van dit gewas zijn weergegeven in Tabel 1. Uit officiële cijfers van FAOSTAT blijkt dat China veruit de grootste producent is van sla met 15,2 miljoen ton productie per jaar. Op de 2de plaats komen de Verenigde Staten met een productie
van ongeveer 3,8 miljoen ton per jaar. India, Spanje en Italië vervolledigen de top 5. In dezelfde periode werd in België 53.370 ton geproduceerd (FAOSTAT, 2017).
Tabel 1 – Sla productie wereldwijd. Cijfers zijn gebaseerd op de productie in 2017 (FAOSTAT, 2017). Productie (miljoen ton/jaar)
China 15,16 Verenigde Staten 3,84 India 1,09 Spanje 0,97 Italië 0,74 België 0,053
In 2017 bedroeg het Belgische areaal groenteteelt ongeveer 50.000 hectare (3,76% van het totale Belgische landbouwareaal). In Vlaanderen nam de teelt van groenten volgens het Vlaams landbouwrapport een oppervlakte in van 31.956 hectare (5,2% van de totale Vlaamse landbouwoppervlakte) (Platteau et al., 2018). Ongeveer 97% van deze oppervlakte was bestemd voor de teelt van groenten in openlucht. De voornaamste teelten van groenten zijn wortelen, bloemkolen, prei, erwten, uien, spruiten en spinazie. Slechts 3% van het groenten areaal in Vlaanderen is bestemd voor de teelt van groenten onder glas. Tomaten en kropsla nemen daar met respectievelijk 48% en 15% de grootste oppervlakte in. Groenten worden vaak in verschillende rondes geteeld waardoor het productiecijfer per hectare hoger zal liggen in vergelijking met andere teelten. De cijfers uit het Vlaams landbouwrapport houden enkel rekening met het areaal dat werd ingenomen voor een welbepaalde teelt (Platteau et al., 2018).
In Vlaanderen wordt kropsla voornamelijk onder glas geteeld (155 ha). Slechts een klein deel (54 ha) wordt in openlucht geteeld. Als men de teeltrotaties zou optellen is er per groeiseizoen sprake van 790 ha kropsla (Statbel, 2017). Naast kropsla wordt ook een klein areaal veldsla geteeld (33 ha, uitsluitend onder glas). Verder worden in Vlaanderen ook alternatieve slasoorten geteeld zoals Lollo Bionda, Lollo Rossa, eikenbladsla, multicolor, rucola en ijsbergsla.
De slateelt in Vlaanderen is onderhevig aan verschillende ziektes en plagen. Enkele belangrijke problemen zijn zwartrot veroorzaakt door Rhizoctonia solani, aantasting door Botrytis cinerea en valse meeldauw veroorzaakt door Bremia lactucae. De meest voorkomende plaag in de slateelt is de bruin-groene slaluis of Nasonivia ribisnigri (Proeftuinnieuws, 2019). De slateelt wordt sinds 2014 bedreigd door een nieuwe ziekte, namelijk de Fusarium-verwelkingsziekte veroorzaakt door Fusarium
oxysporum f. sp. lactucae (Fol). Deze ziekte zorgt voor grote problemen in de Vlaamse slateelt omdat
er geen bestrijdingsmiddelen beschikbaar zijn. Er wordt volop gezocht naar methoden om deze ziekte terug te dringen. Biologische controle kan hierin een belangrijke rol spelen.
4 2.1.2 Bedreiging door Fusarium oxysporum
Beschrijving pathogeen
Fusarium oxysporum forma specialis lactucae (Fol) werd voor het eerst beschreven in 1955 in Japan.
Binnen een forma specialis kunnen verschillende fysio’s of races voorkomen. Voor Fol zijn ondertussen vier verschillende fysio’s beschreven. Fysio 1 tot en met 3 werden in Japan ontdekt. Van daaruit verspreidde de pathogeen zich verder naar verschillende plaatsen in de wereld. Deze verspreiding verliep voornamelijk via gecontamineerd zaad (Garibaldi et al., 2004). In 2002 werd fysio 1 in Europa aangetroffen (Italië en Duitsland) (Pintore et al., 2017). Enkele jaren later (2014) werd Fol ook in Nederland en België gevonden. Volgens onderzoek wijkt de fysio uit Nederland af van de drie eerder beschreven fysio’s voor sla (Gilardi et al., 2017). Deze fysio werd ondertussen als fysio 4 geclassificeerd. Ook in Vlaanderen werd fysio 4 al aangetroffen, wat wijst op een snelle verspreiding (Claerbout et al., 2018). Fysio 4 zorgt in Nederland en België op dit moment voor grote problemen omdat er zeer weinig bestrijdingstechnieken gekend zijn om deze fysio te controleren. Volgens een hypothese van Gilardi et al. (2017) is fysio 4 ontstaan in Nederland door contact van fysio 1 met een niet-pathogene Fusarium stam. Op deze manier kon een nieuwe fysio ontstaan die goed aangepast is aan de klimatologische omstandigheden in België en Nederland. Zo vormen koudere temperaturen niet langer een probleem voor de overleving van de schimmel. Daarnaast zou de fysio ook ontstaan kunnen zijn door een hoge selectiedruk in de plantenveredeling. Verdere studies zijn nodig om het ontstaan van deze nieuwe fysio te verklaren.
Ontwikkeling en verspreiding van de ziekte
F. oxysporum bij sla plant zich enkel ongeslachtelijk voort. Hierbij worden de volgende aseksuele
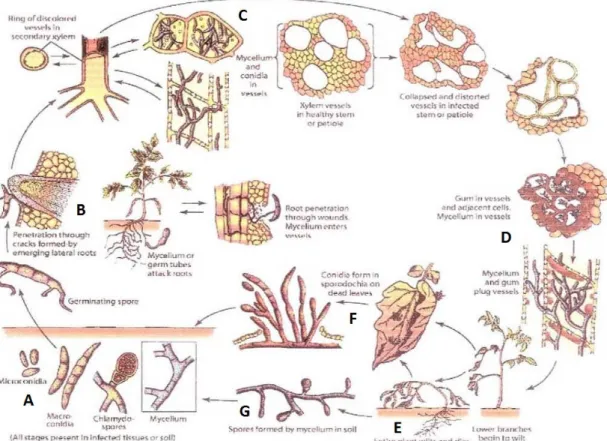
sporen gevormd: microconidia, macroconidia en chlamydosporen (Agrios, 2005). Microconidiën zijn 1- tot 2-cellig en worden geproduceerd door het mycelium. Ze kunnen later zelf kiemen tot nieuw mycelium. Macroconidiën zijn 2- tot meercellig, worden gevormd op een sporodochium en zijn eenvoudig te herkennen aan hun typische halve maan vorm. Chlamydosporen zijn dikwandige sporen die in ongunstige omstandigheden zoals droogte of hitte gevormd worden. Deze sporen dienen als overlevingsstructuur en kunnen meerdere jaren in de bodem overleven (Ford et al., 1970). De verschillende sporen worden weergegeven in Figuur 1-A.
In een substraat met mycelium of sporen worden kiembuizen gevormd in de nabijheid van slaplanten. Deze kiembuizen penetreren de wortels via de worteltip of via wondjes ontstaan door de groei van laterale wortels (Figuur 1-B). De schimmel begeeft zich vervolgens naar het vaatbundelweefsel en zal zich daar vestigen (Agrios, 2005). Via de xyleemvaten worden sporen verder doorheen de plant verspreid (Figuur 1-C). De verspreiding via sporen in de sapstroom is een vorm van passieve verspreiding. Fusarium kan zich ook op een actieve manier verspreiden door het vormen van hyfen en mycelium.
Om de infectie van de schimmel af te stoppen zal de plant reageren door pectineuze bestandsdelen en ‘wound gums’ af te scheiden (Figuur 1-D). Deze elementen komen in het xyleemweefsel terecht en dragen op die manier ongewild bij aan de verstopping. De geïnfecteerde cellen ondergaan ook hyperplasie en hypertrofie. Uiteindelijk zullen de vaatbundels verstopt raken met microconidiën, mycelium en polysachariden. De normale doorstroming van water doorheen het xyleem komt in het gedrang waardoor in de bladeren een watertekort ontstaat met verwelking tot gevolg (Figuur 1-E). Eens volledig afgestorven worden aan het oppervlak van de plant door de pathogeen opnieuw sporen gevormd (Figuur 1-F). Deze kunnen via water en wind worden overgedragen naar naburige planten. Daarnaast kunnen ook nieuwe sporen gevormd worden uit het mycelium dat overgebleven is in de bodem (Figuur 1-G).
5
Figuur 1 – Levenscyclus van Fusarium oxysporum f.sp. lycopersici op tomaat. De cyclus op tomaat is zeer gelijkaardig als deze op sla en verloopt als volgt: (A) van links naar rechts: microconidiën, macroconidiën, chlamydosporen en mycelium, (B) penetratie van gekiemde sporen of mycelium in laterale wortels via worteltip of wondjes, (C) vestiging en aantasting door mycelium en conidiën in vaatbundels, (D) vorming van wound gums door de plant, (E) verwelking door verstopping van xyleem, (F) vorming van nieuwe conidiën op dode bladeren en (G) vorming van sporen uit mycelium uit de bodem (Agrios, 2005).
De verspreiding van Fol kan op verschillende manieren gebeuren. Over het algemeen wordt aangenomen dat zaadoverdracht bijdraagt aan de verspreiding over grotere afstanden. De verspreiding op korte afstanden gebeurt eerder via ziek plantmateriaal, plantenresten, waterdruppels en verstuiving in de lucht. Daarnaast kan ook besmette grond een bron zijn van infectie. Via aarde die aan werktuigen blijft kleven, kan Fusarium overgebracht worden tussen percelen (Agrios, 2005). Symptomen van een infectie met Fol worden weergegeven in Figuur 2. De plant vertoont duidelijke verwelkingssymptomen (Figuur 2-A). Verstopping van de xyleemvaten is zichtbaar als bruinverkleuring (Figuur 2-B). De plant zal uiteindelijk volledig afsterven ten gevolge van de Fusarium infectie.
Figuur 2 – Typische symptomen van de Fusarium verwelking bij sla. (A) Verwelkte bladeren ten gevolge van de verstopte xyleemvaten en (B) bruinverkleuring door aanwezigheid Fol en verstopping xyleemvaten (EPPO, 2019).
6
2.1.3 Bestaande bestrijdingstechnieken tegen Fusarium oxysporum Teelt technisch
Voorlopig zijn hygiënemaatregelen de enige echte oplossing om besmetting en verdere verspreiding van Fol te voorkomen. In Nederland werd een hygiëneprotocol opgesteld door ‘Groen Agro Control’ in samenwerking met ‘LTO Glaskracht Nederland’ om verspreiding van Fusarium tussen landbouwbedrijven te voorkomen. Op basis van dit verslag werd ook in België een protocol uitgewerkt. Vooral de grondtemperatuur heeft een grote invloed op de groei. Hoe hoger de temperatuur van de bodem, hoe makkelijker Fol sla kan infecteren (Scott et al., 2010).
Een andere mogelijke strategie voor de beperking van Fol aantasting is de overstap naar hydrocultuur. Door verschillende problemen met bodem gebonden pathogenen vindt in de slateelt een transitie plaats naar hydrocultuur. In deze systemen kan de infectiedruk verlaagd worden door onder andere een ontsmetting van het recirculatiewater en het gebruik van ziektevrije substraten. Er is geen garantie dat Fusarium niet zal optreden want het is een water minnende schimmel die zich zeer snel kan verspreiden. Tot op heden zijn er geen meldingen van natuurlijke Fol aantastingen in hydrocultuur systemen in België en Nederland. Het nadeel van dergelijke systemen zijn de hoge investeringskosten. Omdat hydrocultuur een intensief systeem is kan de infectiedruk van andere pathogenen ook stijgen (Claerbout, 2020). Daarnaast moet de teelt nauwer opgevolgd worden door de aanwezigheid van een kleinere nutriënten- en waterbuffer.
Het gebruik van resistente rassen lijkt een effectieve manier om Fol te onderdrukken. Echter zijn deze rassen enkel geschikt voor vollegrondsteelten en zijn ze minder productief. Voor de teelt van kropsla zijn bijna alle types onder glas min of meer gevoelig voor Fol. De huidige teelttechniek laat dus niet toe om over te schakelen naar volledig of intermediair resistente rassen. Via verdere veredeling kunnen stappen gezet worden naar rassen die zich kunnen verweren in de huidige teelttechniek.
Vruchtafwisseling is minder geschikt om Fol te onderdrukken. Onderzoek toonde aan dat de pathogeen in staat is om andere gewassen te koloniseren. Zo zouden de wortels van tomaat, broccoli, bloemkool en spinazie gekoloniseerd kunnen worden zonder zichtbare symptomen (Scott et al., 2014). Op deze manier blijft de infectiedruk hoog zonder dat de landbouwer er zich van bewust is. Daarnaast kunnen de chlamydosporen van Fol ook zeer lang in de bodem overleven. Sommige sporen kunnen tot wel 30 jaar dormant blijven (Couteaudier & Alabouvette, 1990; Koike et al., 2009). Dit maakt het systeem van vruchtafwisseling ongeschikt om Fol te onderdrukken.
Biologische controle
Door de vele negatieve effecten geassocieerd met pesticidegebruik wordt volop gezocht naar alternatieve controlemechanismen in de landbouw. Biocontrole organismen zijn een belangrijk voorbeeld van dergelijke alternatieven. Deze organismen kunnen onder andere parasitair zijn voor Fol of in competitie gaan voor nutriënten in het nadeel van de pathogeen. Zo kunnen sommige niet-pathogene Fusarium soorten de verwelkingsziekte reduceren door in competitie te treden met de pathogene soort (Forsyth et al., 2006). Daarnaast kunnen biocontrole organismen ook secundaire metabolieten produceren die toxisch zijn voor de pathogeen en kunnen ze plant resistentie induceren (Alabouvette et al., 2009). Vele van deze potentiële biocontrole organismen zitten nog in de onderzoeksfase en kennen geen commerciële toepassing. Tot op heden zijn de volgende biologische middelen toegelaten in België voor de bestrijding van Fusarium bij sla: PRESTOP (32% Clonostachys
rosea J1446), PRESTOP MIX (26% Clonostachys rosea J1446) en SERENADE ASO (13.96 g/l Bacillus amyloliquefaciens QST 713) (Fytoweb, 2019).
7 Chemische middelen
Een van de laatste opties in een gezond gewasbeschermingsmodel is het gebruik van chemische gewasbeschermingsmiddelen. Voor de bestrijding van Fusarium in sla zijn op dit moment geen middelen toegelaten in België. Vaak zijn grote bedrijven niet geïnteresseerd in relatief kleine gewassen (Spadaro & Gullino, 2005). Daarnaast hebben fungiciden in het verleden weinig succes gehad tegen
Fusarium. De normen van gewasbeschermingsmiddelen op vlak van milieu en gezondheid worden ook
steeds strenger. Bedrijven zullen minder investeren in de ontwikkeling van nieuwe pesticiden omdat aan te veel voorwaarden moet voldaan zijn. De ontwikkeling is te duur om rendabel te zijn. Ondertussen zijn ook vele bestaande producten verboden waardoor gezocht moet worden naar alternatieven.
2.2 Boon – Rhizoctonia solani & Botrytis cinerea
2.2.1 Inleiding bonenteelt
De gewone boon of Phaseolus vulgaris L. behoort tot de Fabaceae of vlinderbloemenfamilie. Volgens gegevens van FAOSTAT uit 2017 is China veruit de grootste producent van P. vulgaris. België heeft een productie van ongeveer 0,12 miljoen ton per jaar (Tabel 2).
Tabel 2 – Productiecijfers van P. vulgaris gebaseerd op de productie in 2017 (FAOSTAT, 2017)
België is vooral gekend voor de verwerking van diepvriesbonen. De bonen die hier geteeld worden zijn dan ook hoofdzakelijk bestemd voor de verwerkende industrie. Deze verwerkende industrie heeft een groot aandeel aan export waardoor bedrijven zelfs bonen aankopen in andere landen. In 2017 werd in België ongeveer 5.450 ha aan bonen geteeld (Statbel, 2017). Het grootse deel van deze teelt is gesitueerd in Wallonië, met een oppervlakte van 3.450 ha. In Vlaanderen wordt ongeveer 2.000 hectare geteeld. Een verwaarloosbaar deel van de bonen wordt onder glas geteeld. Van de geteelde bonen zijn de prinsessenbonen en de zeer fijne ‘haricots verts” het meest gekend. Bonen behoren tot de vlinderbloemigen waardoor ze in staat zijn om, in symbiose met een aantal bacteriën, stikstof uit de atmosfeer te fixeren. Bonen en bij uitbreiding alle vlinderbloemigen zijn om deze redenen zeer interessante gewassen.
De meest belangrijke ziektes op boon zijn schimmelziektes. Deze ziektes zijn vaak ook het moeilijkst om te bestrijden. Voetziektes worden bijvoorbeeld veroorzaakt door Rhizoctonia solani, Fusarium
solani en Fusarium oxysporum. Hierbij zal een roodbruine verkleuring te zien zijn aan het begin van het
wortelstelsel (Inagro, 2019). De planten kunnen hierdoor volledig afsterven. Een andere belangrijke ziekte is peulrot veroorzaakt door Botrytis cinerea. Op de peulen zal een grijs mycelium groeien waardoor de peulen uiteindelijk zullen rotten. Tot slot is sclerotiniarot veroorzaakt door Sclerotinia
sclerotiorum een belangrijke belager in de bonenteelt. Het wit schimmelpluis kan zich razendsnel
verspreiden en de planten afdoden. Een belangrijke plaag op boon is de bonenspintmijt of Tetranychus
urticae. Op het blad zijn witte stippen zichtbaar en bij ernstige schade zullen de bladeren afvallen.
Schade door spintmijten is goed te herkennen aan de productie van spinseldraden. In deze thesis wordt verder ingegaan op R. solani en B. cinerea als belangrijke pathogenen.
Productie (miljoen ton/jaar)
China 19,29 Indonesië 0,92 India 0,68 Turkije 0,63 Thailand 0,31 België 0,12
8 2.2.2 Bedreiging door Rhizoctonia solani
Beschrijving pathogeen
R. solani veroorzaakt zwartrot bij verschillende waardplanten. Het is een plant pathogene schimmel
met een necrotische levensstijl. De schimmel vormt ruststructuren (sclerotiën) die jaren in de bodem kunnen overleven. R. solani bestaat uit een soortencomplex dat onderverdeeld is in anastomosegroepen (AG’s). Een anastomosegroep identificeert een groep van schimmels van dezelfde soort die in staat zijn tot fusie van hun hyfen en het uitwisselen van genetisch materiaal. R. solani bevat meerdere kernen per cel en wordt onderverdeeld in AG1 tot AG13. Niet alle anastomosegroepen zijn pathogeen (Garcia et al., 2006). Binnen deze onderverdeling bestaan nog verdere subgroepen (Agrios, 2005). Bij boon werden volgende anastomosegroepen vaak teruggevonden: AG-1-IA, AG-1-IB, AG-2-1, AG-2-2, AG-4 en AG-5 (Muyolo et al., 1993; Echavez-Badel et al., 2000; Garcia et al., 2006). Lang werd aangenomen dat de schimmel zich enkel ongeslachtelijk kon vermeerderen, maar onlangs is ook de productie van basidiosporen beschreven. Het voorkomen van basidioporen is wel afhankelijk van anastomosegroep tot anastomosegroep. De rol van deze sporen bij de ziekteontwikkeling of verspreiding van de schimmel is echter nog niet volledig uitgeklaard.
Ontwikkeling en verspreiding ziekte
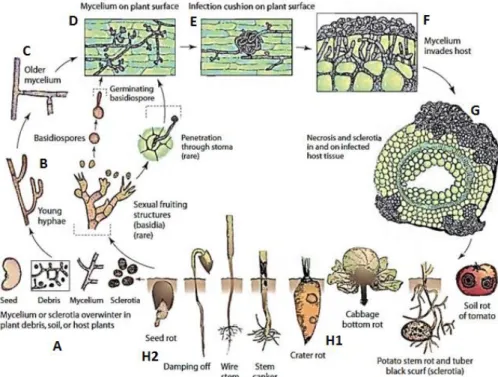
De levenscyclus van R. solani wordt weergegeven in Figuur 3. De schimmel overwintert in de bodem of op achtergebleven plantmateriaal via mycelium en scleroten (3-A). De jonge hyfen groeien uit tot ouder mycelium dat zich vertakt in hoeken van 90 graden (3-B-C). Via infectiekussens zal het mycelium het oppervlak van de plant koloniseren en binnendringen (3-D-F). De schimmel kan de plant binnendringen via zaden, wortels, stengels en bladeren (Abawi et al., 1985). Uiteindelijk treedt necrose op en zullen scleroten gevormd worden (3-G) (Agrios, 2005). De schimmel kan verder verspreid worden via regen, water en machines. Ook besmet plantenmateriaal is een vorm van verspreiding.
Figuur 3 - Levenscyclus van Rhizoctonia solani. (A) Het overleven op plantenmateriaal en in de bodem als mycelium en scleroten, (B) vorming van jonge hyfen, (C) vorming van oudere hyfen (90 graden), (D) koloniseren van het plantenoppervlak, (E) vorming van een infectiekussen, (F) binnendringen van het plantenweefsel, (G) necrosis en het vormen van scleroten op en in het weefsel, (H1) wortelrot en (H2) uitval van jonge kiemplanten (Agrios, 2005).
9
De verschillende symptomen op een bonenplant na infectie met R. solani zijn terug te vinden in Figuur 4. Op de Figuur is zowel een infectie via het blad als een infectie via de wortels door R. solani te zien. De gezonde bladeren en wortels zijn weergegeven ter vergelijking.
Figuur 4 – Symptomen van R.solani infectie op boon waarbij (A) een gezond bonenblad, (B) infectie met R.solani AG2-2 na vijf dagen, (C) een gezonde wortel van boon en (D) infectie met R.solani AG 4-HGI na zeven dagen (Ma, 2016).
2.2.3 Bestaande bestrijdingstechnieken tegen Rhizoctonia solani Teelt technisch
Een goede bedrijfshygiëne speelt een belangrijke rol in de verdere verspreiding van R. solani naar andere landbouwpercelen. Daarnaast kan een efficiënte onkruidbestrijding de Rhizoctonia populatie helpen in te perken. Deze onkruiden kunnen namelijk de schimmel in stand houden en makkelijk overdragen naar het gewas. Ook voldoende teeltrotatie kan een positief effect hebben. Opnieuw is het kiezen van een resistente of tolerante cultivar van groot belang. Volledige resistentie is voorlopig niet mogelijk maar via plantenveredeling wordt bijvoorbeeld in suikerbiet al volop gezocht naar tolerante cultivars (SES-VanderHaves, 2019). Het nadeel van resistente cultivars is de lagere productiviteit en het feit dat bij grote infectiedruk nog steeds veel uitval plaatsvindt. Daarnaast zijn jonge kiemplantjes ook gevoeliger voor uitval door R. solani, hierdoor kan zelfs bij tolerante variëteiten een hoog uitvalcijfer voorkomen.
Biologische controle
Trichoderma harzianum toont veel potentieel in de biologische bestrijding van R. solani bij bonen. Trichoderma is een schimmel die frequent voorkomt in bodems. Al lang is geweten dat Trichoderma
soorten antagonistische eigenschappen vertonen ten opzichte van meerdere economisch belangrijke pathogenen (Howell, 2003). Trichoderma soorten hebben verschillende werkingsmechanismen als mycoparasitisme, antibiosis, competitie, enzymactiviteit en het induceren van plantresistentie. Zowel onder glas als in volle grond werd aangetoond dat T. harzianum in staat was om bonen te beschermen tegen R. solani (Elad et al., 1980). Daarnaast vertonen sommige bacteriën ook potentieel om als biocontrole organismen ingezet te worden. Zo werd gevonden dat Bacillus subtilis RB14 een belangrijke rol speelde in het onderdrukken van uitval van tomaten kiemplanten door R. solani (Asaka & Shoda, 1996).
R. solani kan ook bestreden worden door gebruik te maken van biofumigatie via een groenbemester.
Verschillende Brassica soorten zoals onder andere Brassica juncea bevatten hoge gehaltes aan glucosinolaten. Deze glucosinolaten zullen na inwerking in de bodem omgezet worden tot isothiocyanaten die op hun beurt een toxische werking hebben tegen R. solani (Charron et al., 1999).
10 Chemische controle
R. solani bestaat zoals hierboven vermeld uit meerdere anastomosegroepen. Eenzelfde fungicide is
niet werkzaam tegen alle anastomosegroepen (Campion et al., 2003). Vertegenwoordigers van verschillende anastomosegroepen zijn sterk, gemiddeld of niet sensitief voor een bepaald product. Op die manier moet per anastomosegroep een werkzaam fungicide gevonden worden. Tot op heden zijn geen fungiciden toegelaten voor de Rhizoctonia bestrijding in de bonenteelt in België (Fytoweb, 2020). Een effectieve stof in het verleden was de bodemontsmetter methylbromide. Het gebruik van deze stof is sinds 2005 verboden omdat het de ozonlaag aantastte (EU Pesticides Database, 2020).
2.2.4 Bedreiging door Botrytis cinerea Beschrijving pathogeen
Ziekten veroorzaakt door B. cinerea zijn wellicht de meest verspreide ziekten van groenten, sierplanten en vruchten in de wereld. Botrytis kan een enorme verscheidenheid aan planten infecteren. Het zeer brede waardplantspectrum maakt van B. cinerea een moeilijke pathogeen om te bestrijden. Botrytis kan verschillende delen van planten aantasten gaande van bloesems, vruchten tot wortelen (Agrios, 2005). B. cinerea is een necrotroof, het zal stoffen uitscheiden waardoor de plantencellen afsterven. Deze afgestorven plantencellen kan de schimmel dan als voeding gebruiken (Koppert, 2020). De schimmel heeft nood aan koel en vochtig weer voor optimale groei en produceert een typisch grijs mycelium op de aangetaste weefsels. Wat het meeste voorkomt zijn grijsbruine plekken op de plaats van infectie. Botrytis is een schimmel die heel vaak voorkomt als secundaire infectie. Het zal dus planten infecteren die al wat verzwakt zijn door andere pathogenen. Zo kan de schimmel bijvoorbeeld makkelijk infecteren via wondjes (Koppert, 2020).
Ontwikkeling en verspreiding ziekte
De levenscyclus van Botrytis wordt weergegeven in Figuur 5. Botrytis overwintert als mycelium in plantenresten of als scleroten (5-A) (Agrios, 2005). De scleroten kunnen kiemen en mycelium vormen om direct te infecteren (5-B), of ze kunnen apothecia met ascosporen vormen (niet te zien op de Figuur). Het mycelium zal uiteindelijk conidioforen aanmaken met de productie van conidiosporen in karakteristieke trossen (5-C). Deze conidiën kunnen het weefsel binnendringen via wonden (5-D), of via productie van mycelium op bloembladeren, dode bladeren, … (5-E) (Agrios, 2005). Na infectie zullen de cellen uit elkaar vallen en afsterven, het weefsel wordt zacht en begint te rotten (5-F). Op het geïnfecteerde weefsel zullen opnieuw conidioforen en conidiosporen aangemaakt worden die in staat zijn om andere planten te infecteren (5-G). B. cinerea kan op vele verschillende plantenresten overleven, waardoor de ziekte moeilijk te bestrijden is (Agrios, 2005).
Verspreiding komt veelvuldig voor na plant-plant contact. Besmette bladeren die in contact komen met andere delen van de plant zullen ook geïnfecteerd raken. Daarnaast hebben nabijgelegen planten ook een hoog risico om geïnfecteerd te raken. Ook via de lucht kunnen de sporen van Botrytis eenvoudig verspreid worden. Vooral onder vochtige omstandigheden is de overdracht van sporen het grootste (Agrios, 2005).
11
Figuur 5 – Levenscyclus van Botrytis cinerea. (A) Het overwinteren als mycelium, (B) productie van mycelium uit scleroten, (C) vorming van conidioforen met conidioporen, (D) infectie via wonden, (E) infectie via mycelium, (F) rotten van weefsel en (G) vorming van nieuwe conidiosporen (Agrios,2005).
De symptomen van B. cinerea op boon zijn terug te vinden in Figuur 6. Er is een kunstmatige infectie op het blad te zien met een sporenoplossing. Er vormt zich een duidelijke roodachtige laesie op het blad. Deze symptomen werden na drie dagen vastgesteld. In Figuur 6-B is een peul te zien die volledig omgeven is met grijs mycelium van Botrytis cinerea.
Figuur 6 – Symptomen op boon na infectie van B. cinerea (A) op blad met sporenoplossing (eigen foto) en (B) op vrucht met vorming van grijs mycelium (bron: NYSIPM Image Gallery)
12
2.2.5 Bestaande bestrijdingstechnieken tegen Botrytis cinerea Teelt technisch
Om tot een effectieve bestrijding te komen is het belangrijk om geïnfecteerde planten en plantenresten zo snel mogelijk te verwijderen (Williamson et al., 2007). De sporen kunnen zich namelijk eenvoudig via de lucht verspreiden. Door het wegnemen van het geïnfecteerd plantenmateriaal slaagt Botrytis er minder goed in om snel van de ene plant naar de andere over te gaan. Aangezien Botrytis een voorkeur heeft voor koudere omgevingstemperaturen is het een goede strategie om serres voldoende te verluchten en te verwarmen (Agrios, 2005). Een hoge luchtvochtigheid dient vermeden te worden. Het is ook van belang om andere plagen zoveel mogelijk te vermijden omdat Botrytis in zo’n gevallen voor secundaire schade zorgt. Overmatig gebruik van stikstof zorgt voor een versnelde vegetatieve groei, wat kan leiden tot een verhoogd risico op Botrytis en andere infecties (Williamson et al., 2007). Voldoende afstand tussen de planten kan de verspreiding ook terugdringen. Uiteraard is het onderhouden van een goede bedrijfshygiëne ook hier essentieel om infectie en verspreiding te vermijden. Ook veredelen van verschillende gewassen is een optie, maar dit blijkt moeilijk omdat B. cinerea snel resistentie doorbreekt (Williamson et al., 2007). Daarnaast zorgt het brede waardplantspectrum ervoor dat de schimmel makkelijk kan overleven op andere gewassen. In de meeste gewassen is veredelen moeilijk en onsuccesvol gebleken. In tomaat is vaak onderzocht of resistentie kan ingekruist worden via wilde varianten uit hun oorsprongsgebied (ten Have et al., 2007; Smith et al., 2014). Tot op heden is nog geen cultivar van tomaat beschikbaar met resistentie tegen Botrytis cinerea. Bepaalde cultivars zijn wel toleranter dan andere.
Biologische controle
In een correct IPM systeem komt biologische controle boven chemische controle. Reeds vroeg werd gerapporteerd dat het gebruik van microbiële antagonisten effectief was om Botrytis ziekten te reduceren (Williamson et al., 2007). Verschillende bacteriën als Streptomyces griseoviridis, Bacillus
subtilis en Pseudomonas syringae vertonen potentieel om aan biocontrole te doen. Daarnaast is er
veel aandacht voor het gebruik van antagonistische schimmels zoals Trichoderma sp. Zo werd aangetoond dat Trichoderma harzianum T39 in staat is om resistentie te induceren om zo de plant te beschermen. Bij de teelt van tomaat, sla, peper, tabak en boon zorgde applicatie van T. harzianum T39 voor 25 tot 100% reductie van Botrytis symptomen door uitstel of onderdrukking van de laesies. In boon had T. harzianum T39 hetzelfde effect als de rhizobacteriën Pseudomonas aeruginosa KMPCH, een modelbacterie voor het induceren van systemische resistentie (De Meyer et al., 1998).
Chemische controle
De laatste optie voor de bestrijding van Botrytis zijn chemische middelen. Het gebruik van fungiciden bleek in het verleden slechts gedeeltelijk succesvol, omdat Botrytis zeer snel resistenties kon doorbreken (Leroux et al., 2002). Op dit moment zijn er geen chemische middelen meer toegelaten in België ter bestrijding van de schimmel in de bonenteelt (Fytoweb, 2020). Omdat bij herhaalde toepassingen te snel resistentie optrad, werd vroeger gebruik gemaakt van vijf verschillende types fungiciden (De Meyer et al., 1998). Uiteindelijk bleken al deze stoffen schadelijk voor mens en milieu waardoor ze van de markt verdwenen zijn.
13
2.3 CLP producerende bacteriën als biocontrole oplossing
2.3.1 Verschuiving van chemische naar biologische bestrijding
Doorheen de geschiedenis van de landbouw werden pesticiden steeds belangrijker om hogere productiecijfers te behalen. Rachel Carson was één van de eersten die kritiek had op de conventionele pesticiden en hun impact op de mens en het milieu. In haar boek ‘Silent Spring’ uit 1962 is ze vernietigend over DDT en andere stoffen door hun impact op het milieu. Tot op heden is er een hevige discussie over de impact van landbouw op het milieu. Hoewel haar werk enorm veel kritiek kreeg, zette dit een gedachteverandering op gang. Vanaf de jaren 2000 werd haar werk weer actueel. Door vele milieu-impactstudies werden heel wat schadelijke producten uit de handel genomen. Pesticiden waren onder andere verantwoordelijk voor bodemverontreiniging en contaminatie van grondwater en waterlopen (Stehle & Schulz, 2015). Nog belangrijker was het feit dat vele van deze producten ook de gezondheid van de mens in gevaar brachten (Di Renzo et al., 2015). Het is voor de wetenschap dan ook de uitdaging om op zoek te gaan naar bestrijdingstechnieken met een verminderde impact op het milieu.
Biologische landbouw toont potentieel om de impact op het milieu te verminderen. Een rechtstreeks en duurzaam voordeel is het verhogen van organische koolstof in de bodem (Pimentel et al., 2005). Ook in de biologische landbouw worden pesticiden gebruikt, zogenoemde ‘natuurlijke pesticiden’. De stoffen die gebruikt worden zijn afkomstig van natuurlijke bronnen. Helaas kunnen ook deze stoffen enorm schadelijk zijn voor het milieu. Stoffen als kopersulfaat zijn bijvoorbeeld meer toxisch dan bepaalde synthetische producten (Lewis et al., 2016). Over de voor- en nadelen van biologische landbouw wordt tot op heden stevig gedebatteerd. Er is onder andere kritiek op het landgebruik, pesticide gebruik en energieverbruik bij biologische landbouw.
Landbouwsystemen waarbij gebruik gemaakt wordt van geïntegreerde controle hebben veel potentieel in het reduceren van de negatieve impact op mens en milieu. De minst schadelijke oplossingen worden naar voor gebracht, waarbij preventie een grote rol speelt. Het gebruik van biologische middelen komt in een Integrated Pest Management (IPM) strategie boven het gebruik van chemische middelen. Biocontrole is een belangrijke tool in de IPM strategie. Het maakt gebruik van verschillende organismen om ziekten, plagen en onkruiden te controleren (Höfte & Altier, 2010).
14 2.3.2 Cyclische lipopeptiden
Structuur
Lipopeptiden (LP) krijgen veel aandacht wegens hun antimicrobiële, cytotoxische en surfactant eigenschappen. Ze worden geproduceerd door schimmels als Aspergillus, en door meerdere bacteriën zoals Streptomyces, Pseudomonas en Bacillus (Raaijmakers et al., 2010). De laatste jaren is er veel aandacht voor cyclische lipopeptiden (CLP’s) geproduceerd door Pseudomonas en Bacillus species als secundaire metabolieten.
CLP’s zijn veelzijdige moleculen die bestaan uit een hydrofiel peptidedeel gekoppeld aan een hydrofoob vetzuurresidu. Het peptidedeel is cyclisch en vormt een lactone ring tussen twee aminozuren in de peptideketen (Raaijmakers et al., 2006). CLP’s worden niet-ribosomaal geproduceerd op grote multifunctionele niet-ribosomale peptide synthetasen (NRPS) via een ‘thiotemplate’ proces (Raaijmakers et al, 2006). Deze synthetases zijn niet enkel betrokken bij de productie van CLP’s, ook de productie van andere belangrijke peptides zoals penicilline loopt via deze pathway. Het NRPS-systeem bezit een structuur die uit modules bestaat waarbij elke module een stap is in de incorporatie en modificatie van één aminozuur. Daarnaast kunnen deze modules ook meerdere niet-proteïne structuren als D-aminozuren, carboxylzuren en vetzuren incorporeren (Gewolb, 2002; Sieber & Marahiel, 2005). Voor een meer gedetailleerde uitleg over dit systeem wordt verwezen naar het werk van Gewolb uit 2002. De diversiteit aan CLP’s is te wijten aan de lengte en configuratie van de vetzuurstaart en peptideketen. CLP’s kunnen interageren met celmembranen van verschillende organismen zoals schimmels, bacteriën en planten (Raaijmakers et al., 2010).
Biologische functie
Via membraaninteracties zijn CLP’s in staat om eigenschappen van bepaalde membranen te wijzigen. Op die manier kan bijvoorbeeld een pathogeen onderdrukt worden. Zo werd van verschillende CLP’s een antibiotische werking beschreven tegen tal van bacteriën en schimmels (Raaijmakers et al., 2006). Verder vertonen sommige CLP’s antivirale activiteit. CLP’s kunnen ook een rol hebben als virulentiefactor bij plant pathogene bacteriën (Raaijmakers et al., 2006). Ze hebben bijvoorbeeld een invloed bij de formatie van openingen in de celmembraan met cellyse tot gevolg.
Naast directe antagonisme kunnen CLP’s ook bepaalde immuunresponsen induceren bij planten. Er zijn verschillende manieren waarop een afweerreactie door de plant tot stand kan komen. Een plant kan immuniteit verworven hebben na contact met een pathogeen. De pathogeen zal een reactie opwekken waardoor de defensiemechanismen van de plant op een hogere intensiteit zullen werken. Deze soort van immuniteit wordt ‘Systemic Acquired Resistance (SAR)’ genoemd. SAR wordt gekarakteriseerd door een verhoging van het salicylzuur in de plant (van Loon et al., 1998). Ook niet-pathogene rhizobacteriën kunnen systemische resistentie induceren, ‘Induced Systemic Resistance (ISR)’ genoemd. ISR wordt over het algemeen opgewekt door wortel koloniserende bacteriën die signaal transductiesystemen afhankelijk van de plantenhormonen jasmijnzuur en ethyleen opwekken (Pieterse et al., 1998).
15
De ‘microbe associated molecular patterns’ (MAMP’s) en ‘pathogen associated molecular patterns’ (PAMP’s) zijn respectievelijk microbe en pathogeen specifieke molecules die bekend staan als elicitoren (Newman et al., 2013). Deze worden herkend door receptoren die geassocieerd zijn met het celmembraan en kunnen zo de immuniteit van de plant verhogen (Boutrot & Zipfel, 2017). Diverse elicitoren die gesecreteerd worden door plant bevorderende bacteriën kunnen ISR triggeren, waaronder de CLP’s (De Vleesschauwer & Höfte, 2009; Mariutto & Ongena, 2015). Deze elicitoren zijn gelijkaardig aan de MAMP’s en de PAMP’s. Het verschil is dat er nog geen indicaties zijn over specifieke CLP-receptoren. Resultaten met het CLP surfactin, geproduceerd door Bacillus species, geven aan dat het om een lipide gedreven proces gaat. Er is een verstoring ter hoogte van het plasmamembraan waardoor ISR getriggerd wordt (Henry et al., 2011).
Bij Arabidopsis planten waar ISR getriggerd werd, kon een ‘priming’ effect vastgesteld worden in vergelijking met niet geïnduceerde planten. Door het effect werden afweer gerelateerde genen eerder en in hogere mate tot expressie gebracht (Conrath et al., 2002). Naast een verhoogde genexpressie kunnen ook structurele barrières versterkt worden om meer weerwerk te bieden tegen pathogenen.
Arabidopsis planten met geïnduceerde immuniteit door P. fluorescens WCS417r vertoonden op de
plaats van infectie met de valse meeldauw pathogeen Hyaloperonospora arabidopsidis een verhoogde frequentie van callose depositie. De calloseproppen zorgden ervoor dat het binnendringen van de pathogeen stopte (Van der Ent et al., 2009). De callose depositie wordt opgewekt door het plantenhormoon abscisinezuur (Ton et al., 2005).
Tot slot zijn CLP’s belangrijk voor de beweeglijkheid (‘swarming motility’) en de biofilm vorming van bacteriën. Bij mutanten met een deficiëntie in de CLP-productie werd bijvoorbeeld een verminderde beweeglijkheid en een verminderde biofilm vorming opgemerkt (de Bruijn et al., 2007; Li et al., 2013; D'aes et al., 2014). De bewegelijkheid kon weer hersteld worden door de externe toevoeging van CLP’s (D'aes et al., 2014).
16 2.3.3 Pseudomonas biocontrole organismen
Taxonomie
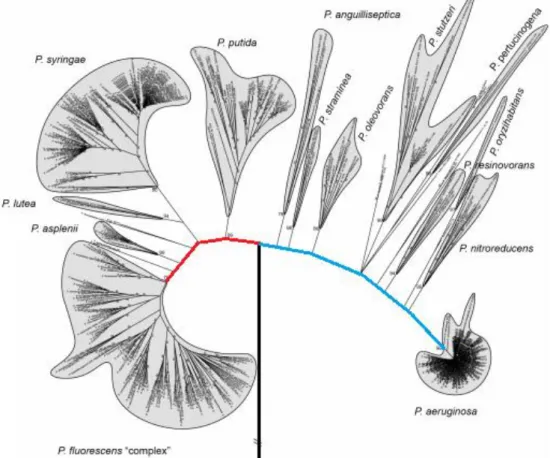
Het genus Pseudomonas bevat ongeveer 220 beschreven species en vormt daarmee de grootste groep van de Gram-negatieve bacteriën (Parte, 2018). Het geslacht Pseudomonas behoort tot de subklasse Proteobacteriën en is gekend voor zijn alomtegenwoordigheid in de omgeving. Pseudomonas bacteriën komen frequent voor in de bodem omdat ze veel verschillende organische componenten kunnen gebruiken als energie- en koolstofbron (Wu et al., 2011). De abundantie hangt af van verschillende factoren zoals het soort plantenweefsel waarop ze leven, het bodemtype, de bodemstructuur, de temperatuur en het vochtgehalte (Gardener, 2007). In de groep van de fluorescente Pseudomonas soorten zijn enkele potentiële biocontrole organismen terug te vinden (Bossis et al., 2000). Fluorescente Pseudomonas bacteriën worden hoofdzakelijk onderverdeeld in twee verschillende lijnen: de Pseudomonas fluorescens lijn en de Pseudomonas aeruginosa lijn (Garrido-Sanz et al., 2016). Een duidelijk overzicht van beide lijnen is gegeven in Figuur 7. De P.
fluorescens lijn bestaat bijvoorbeeld uit vijf groepen: P. putida, P. syringae, P. lutea, P. asplenii en P. fluorescens. Van sommige van de soorten uit deze lijn is geweten dat ze effectieve antagonisten zijn
van bodem gebonden pathogenen (Haas & Defago, 2005). De meeste species leven als saprofyten die voorkomen in water en bodem. Ze hebben interessante kenmerken en worden gebruikt in biotechnologische toepassingen om de groei en gezondheid van planten te bewaren (Höfte & Altier, 2010). Sommige species van de Pseudomonas aeruginosa lijn komen veelvuldig voor en kunnen geassocieerd worden met planten en dieren. P. aeruginosa is beter bekend als de ziekenhuisbacterie. De enorme fenotypische en genetische variabiliteit binnen de fluorescente Pseudomonas soorten heeft ervoor gezorgd dat de taxonomie erg ingewikkeld is (Garrido-Sanz et al., 2016).
Figuur 7 – Overzicht van fluorescente Pseudomonas soorten met de twee hoofdlijnen: P. fluorescens “complex” (rode lijn) en P. aeruginosa (blauwe lijn) (Garrido-Sanz et al., 2016). De gekleurde lijnen zijn zelf aangebracht.
17
De introductie van genoomanalyses zorgt voor een nieuwe kijk op de bacteriële taxonomie. De nieuwe inzichten kunnen gebruikt worden om klaarheid te scheppen in de taxonomische chaos. Via nauwkeurige technieken kunnen soorten beter afgebakend worden. De fylogenie van hogere taxonomische rangen kan afgeleid worden, tot uiteindelijk zelfs op het niveau van de subspecies (Lalucat et al., 2020). In de studie van Lalucat et al. (2020) werd recent de huidige taxonomie van
Pseudomonas herbekeken. De onderzoekers konden onder meer twee belangrijke conclusies maken:
(1) verschillende soorten zijn synoniemen van elkaar en moeten tot één soort benoemd worden; (2) veel stammen die aan bekende soorten zijn toegevoegd, moeten worden voorgesteld als nieuwe soorten binnen het geslacht.
Via een ‘multilocus sequence’ analyse gebaseerd op aaneengeschakelde partiële gensequenties van vier geselecteerde genen (16S rDNA, gyrB, rpoB en rpoD) werd voor 216 Pseudomonas species en subspecies onderzocht in welke mate ze verwant waren aan elkaar (Lalucat et al., 2020). Het genus kon verdeeld worden in drie hoofdlijnen (P. aeruginosa, P. fluorescens en P. pertucinogena). De P.
fluorescens lijn bestond inderdaad uit vijf fylogenetische groepen (P. fluorescens, P. asplenii, P. lutea, P. syringae en P. putida). De P. aeruginosa lijn bestond uit acht verschillende fylogenetische groepen
(P. straminea, P. anguilliseptica, P. oryzihabitans, P. stutzeri, P. oleovorans, P. resinovorans, P.
aeruginosa en P. linyingensis) en het genus Azotobacter. Verschillende auteurs stellen voor om het genus Azotobacter binnen het genus van Pseudomonas te brengen (Rediers et al., 2004; Ozen & Ussery,
2012). De resultaten wezen erop dat de lijn van P. pertucinogena eigenlijk als een verschillend genus kan gezien worden. Extra onderzoek is nodig om tot een nieuwe en meer correcte indeling te komen. Op die manier kan Pseudomonas een modelorganisme worden op vlak van bacteriële taxonomie. Voorkomen en variatie van CLP’s
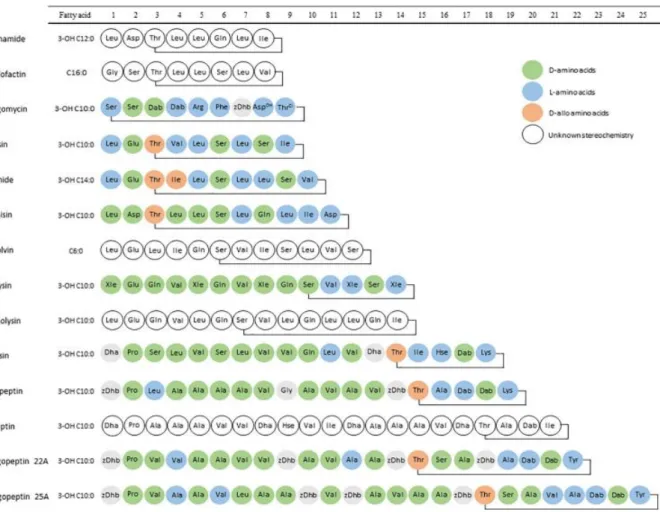
Verschillende Pseudomonas stammen zijn terug te vinden aan het worteloppervlak van planten. Ze komen voor als micro-kolonies of vormen biofilms tussen de epidermale cellen van de wortels. Rezzonico et al. (2007) beschreven dat stammen met potentiële biocontrole eigenschappen 10% uitmaken van alle stammen die te vinden zijn in de rhizosfeer. Dit cijfer werd bekomen door stammen te analyseren uit een groot aantal verschillende bodems met verschillende planten uit alle delen van de wereld. De CLP’s die gevormd worden door de verschillende Pseudomonas species zijn onderverdeeld in meerdere families (Gotze & Stallforth, 2020). Het aantal families is dynamisch en kan nog steeds uitgebreid worden. Ook binnen de verschillende families of groepen kunnen nieuwe vertegenwoordigers geïntroduceerd worden. Volgens het werk van Geudens en Martins uit 2018 kunnen de CLP’s vandaag in 14 verschillende families onderverdeeld worden. De verdeling gebeurt op basis van de chemische structuur van het CLP. De basisstructuur van elke groep is weergegeven in Figuur 8. Enkele grote families konden in het verleden snel gevormd worden. Op deze manier ontstonden de families van de viscosines, de orfamides, de amphisines, de syringomycines, de syringopeptines 22A en de toolasines (Geudens & Martins, 2018). Wanneer nieuwe CLP’s ontdekt worden, zullen deze volgens hun chemische structuur in één van de families geklasseerd worden. Wanneer een CLP geïsoleerd kan worden dat structureel niet bij een bestaande familie hoort, kan een nieuwe familie gedefinieerd worden. Op die manier zijn de groepen van de xantholysines (Li et al., 2013) en de bananamides (Nguyen et al., 2017) recentelijk toegevoegd. Enkele CLP’s zijn zeer moeilijk in een bepaalde familie te plaatsen. Voor deze CLP’s wordt de lengte van de oligopeptide keten gebruikt als criterium voor classificatie. Zo ontstonden de groepen van de entolysines (Vallet-Gely et al., 2010), putisolvines (Kuiper et al., 2004), pseudofactines (Janek et al., 2010), syringopeptines 25A
18
(Ballio et al., 1991), corpeptines (Emanuele et al., 1998) en fuscopeptines (Ballio et al., 1996). Recent werd het CLP cocoyamide A ontdekt, maar deze kon niet aan een bepaalde familie toegevoegd worden (Oni et al., 2019). Voor een vollediger overzicht van de structuren van de verschillende groepen wordt verwezen naar het werk van Geudens en Martins uit 2018. Zeer recent werd een review uitgevoerd over de structuren, families, eigenschappen en biologische functies van CLP’s geproduceerd door
Pseudomonas bacteriën. Er werden slechts enkele kleine wijzigingen gesuggereerd op vlak van de
indeling van de families. Hiervoor wordt verwezen naar het werk van Götze en Stallforth uit 2020.
Figuur 8 – Overzicht van de chemische structuren van de verschillende CLP-families geproduceerd door
Pseudomonas species (Geudens & Martins, 2018). Op dit moment kunnen 14 verschillende families
onderscheiden worden. Legende: D-aminozuur (Groen), L-aminozuur (Blauw), D-allo-aminozuur (Rood) en ongekende stereochemie (Wit).
19 Biocontrole via CLP’s
In Tabel 3 wordt een overzicht gegeven van CLP’s geproduceerd door Pseudomonas species die relevant zijn voor deze thesis. Uiteraard zijn er nog vele andere CLP’s die biocontrole capaciteiten vertonen. Voor een volledig overzicht van de meest relevante CLP’s en hun producenten wordt opnieuw verwezen naar het werk van Geudens en Martins uit 2018 en naar dat van Götze en Stallforth uit 2020.
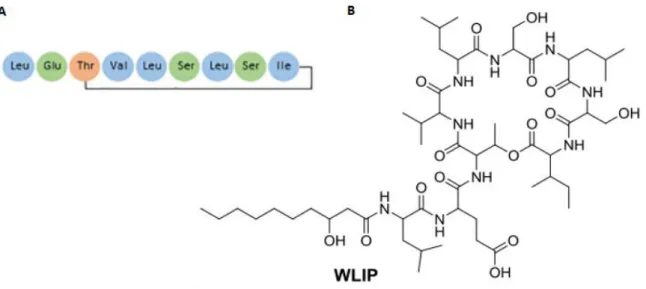
Zoals eerder aangegeven kan de structuur van CLP’s enorm variëren. In Figuur 9 wordt bij wijze van voorbeeld de structuur van WLIP weergegeven (Ma, 2016). WLIP is een CLP uit de viscosine familie en wordt onder andere geproduceerd door Pseudomonas putida RW10S2. WLIP is een lipodepsipeptide met een moleculaire massa van 1125 Da en heeft een peptideketen van negen aminozuren. De lactone ring bevindt zich tussen D-allo-threonine en de C-terminale L-isoleucine. WLIP is zeer gelijkaardig aan het CLP viscosine (Coraiola et al., 2006). De afkorting WLIP staat voor “white-line-inducing principle”. Producenten van WLIP zijn namelijk in staat om, bij aanwezigheid van Pseudomonas tolaasii, een duidelijk zichtbare witte lijn te vormen tussen beide kolonies (Rokni-Zadeh et al., 2012). Dit fenomeen is te wijten aan de reactie tussen WLIP met het CLP tolaasin. Tolaasin is toxisch voor schimmels en behoort tot een andere structurele groep dan WLIP. Het verschijnen van de witte lijn wordt gebruikt als identificatie voor tolaasin producerende bacteriën die schade verrichten in de teelt van champignons. Later werd gevonden dat ook andere CLP’s onderling in staat zijn om een witte lijn te vormen. Zo werd via een test met de mutanten van Pseudomonas sp. CMR12a aangetoond dat orfamides en sessilines eveneens een witte lijn kunnen induceren ten opzichte van elkaar (D’aes et al., 2014).
Figuur 9 - Structuur van het CLP WLIP geproduceerd door onder andere Pseudomonas putida RW10S2. In (A) wordt de samenstelling van de aminozuren gegeven (Geudens & Martins, 2018). In (B) is de volledige structuur gevisualiseerd (Ma, 2016).
Directe antagonisme activiteit van Pseudomonas CLP’s werd reeds in meerdere pathosystemen aangetoond (Tabel 3). Een voorbeeld van directe antagonisme is het gebruik van Pseudomonas sp. CMR12a bij boon. Het werd aangetoond dat het CLP sessiline een rol speelt in het directe antagonisme tegen wortelrot veroorzaakt door R. solani AG 2-2 of R. solani AG 4 HGI (D’aes et al., 2011). Daarnaast toonden experimenten een verhoging van het aantal vertakkingen van de hyfen aan bij beide