DIVERSITEIT VAN DE PHYTOPTHORA
INFESTANS
POPULATIE
IN
VLAANDEREN
GEVOLGEN
VOOR
DE
AGRESSIVITEIT
VAN
DE
AARDAPPELZIEKTE
Alexander Lannoije
Stamnummer: 01508123Promotor(en): Prof. dr. ir. Geert Haesaert, dr. ir. Sofie Landschoot
Tutor: Ing. Kevin Dewitte
Masterproef voorgelegd voor het behalen van de graad in Master of Science in de biowetenschappen: land- en tuinbouwkunde - afstudeerrichting plantaardige en dierlijke productie
DIVERSITEIT VAN DE PHYTOPTHORA
INFESTANS
POPULATIE
IN
VLAANDEREN
GEVOLGEN
VOOR
DE
AGRESSIVITEIT
VAN
DE
AARDAPPELZIEKTE
Alexander Lannoije
Stamnummer: 01508123Promotor(en): Prof. dr. ir. Geert Haesaert, dr. ir. Sofie Landschoot
Tutor: Ing. Kevin Dewitte
Masterproef voorgelegd voor het behalen van de graad in Master of Science in de biowetenschappen: land- en tuinbouwkunde - afstudeerrichting plantaardige en dierlijke productie
“De auteur en de promotor geven toelating deze scriptie voor consultatie beschikbaar te stellen en delen van de scriptie te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze scriptie.”
“The author and the promotor give the permission to use this thesis for consultation and to copy parts of it for personal use. Every other use is subject to the copyright laws, more specifically the source must be extensively specified when using the results from this thesis”
Woord vooraf
Het schrijven van een thesis is een hele uitdaging, zeker voor iemand zoals mij die alles wil perfectioneren en zijn woordkeuze driemaal veranderd vooraleer er uiteindelijk een zin is getypt. Het was een enorm voordeel dat het onderwerp mij interesseerde aangezien wij thuis zelf aardappelen kweken en ik mij daardoor afvroeg waarom deze oömyceet niet beheersbaar is en ik me hierin graag wou verdiepen. Helaas kreeg deze thesis een twist doordat er geen of toch amper P. infestans isolaten werden gevonden in groeiseizoen 2019. Hierdoor moesten er alternatieve proeven opgezet worden.
Vervolgens zou ik graag enkele mensen willen bedanken die mij geholpen hebben bij de realisatie van mijn eindwerk. Zonder hun steun en vertrouwen was het niet mogelijk geweest om mijn thesis tot een goed einde te brengen.
Om te beginnen wil ik mijn twee promotoren bedanken, prof. dr. ir. Geert Haesaert en in het bijzonder dr. ir. Sofie landschoot voor alle feedback en suggesties. Vervolgens wil ik ook mijn tutor, ing. Kevin Dewitte bedanken voor de uitvoering en uitleg bij de proeven.
Daarnaast een woord van dank aan Neda Najdabbasi van de proefhoeven in Bottelare voor de opzet en de hulp bij de proeven.
En als laatste een speciale bedanking aan mijn ouders voor de steun en de mogelijkheid die zij mij hebben gegeven om deze opleiding te volgen. Ik wil ook mijn grootouders, schoonouders en zussen bedanken, alsook in het bijzonder mijn vriendin voor alle lekkere vieruurtjes en motiverende woorden die me wisten verder te gaan als ik het even niet meer zag zitten.
Preambule
Het oorspronkelijk doel was om de P. infestans populatie in kaart te brengen en deze isolaten te gebruiken voor experimenten naar bijvoorbeeld pathogeniciteit toe. In de zomervakantie en tijdens het eerste semester werd er in aardappelpercelen en percelen met aardappelopslag gezocht naar P. infestans isolaten. Door het droge weer was er nauwelijks P. infestans te vinden, waardoor onafhankelijk van de corona crisis, de doelstelling van deze thesis diende bijgestuurd te worden. Als alternatief ging tijdens het tweede semester een selectie van aardappelrassen gescreend worden voor hun gevoeligheid voor verschillende (referentie) P.
infestans genotypen. Verder gingen de infecties zowel op intacte planten als op afgeknipte
bladeren (Detached Leaf Assay) gebeuren. De alternatieve proef kon niet door de student zelf uitgevoerd worden en werd ingekort door maar één referentie genotype te evalueren tegenover verschillende aardappelrassen via een DLA.
‘Deze preambule werd in overleg tussen de student en de promotor opgesteld en door beiden goedgekeurd’
Abstract (Nederlands)
In dit werk werd initieel de diversiteit van P. infestans in Vlaanderen bestudeerd, maar door de voor P. infestans ongunstige weersomstandigheden, te warm en te droog, kon de populatie in Vlaanderen niet in kaart gebracht worden. De alternatieve proeven stonden in het teken van waarom er geen aardappelplaag gevonden werd in de warme zomer van groeiseizoen 2019. In de literatuurstudie werd dieper ingegaan op de geschiedenis, diversiteit, levenscyclus en beheersing van P. infestans. Bij de proeven werd eerst een verklaring gegeven waarom er geen P. infestans isolaten werden gevonden in groeiseizoen 2019. Daarna werd gekeken welke invloed temperatuur had op de myceliumgroei en de sporenkieming bij verschillende isolaten. Om te eindigen werd een proef opgezet waarbij de plaaggevoeligheid werd getest bij verschillende rassen.
Uit de temperatuurproeven werd, op basis van alle isolaten samen, besloten dat myceliumgroeisnelheid maximaal is bij 17,02 °C en 20,24 °C was de optimale temperatuur voor sporenkieming. Hoewel de reactie van de isolaten t.o.v. de verschillende temperaturen gelijkaardig was, waren er toch verschillen. Deze verschillen kunnen een invloed hebben op het voorkomen van bepaalde isolaten in bepaalde regio’s.
Uit de proef omtrent rasgevoeligheid bleek duidelijk dat er significante verschillen zijn tussen de rassen wat betreft de gevoeligheid voor laesie ontwikkeling op de bladeren. Dit is naast opbrengst en kwaliteit een belangrijke factor die moet in rekening gebracht worden bij de keuze van een aardappelras.
Abstract (English)
This work initially studied the diversity of P. infestans in Flanders, but due to the unfavourable weather conditions for P. infestans, too hot and too dry, the population in Flanders could not be mapped. The alternative tests were focused on why no late blight was found in the warm summer of growing season 2019.
In the literature study, the history, diversity, life cycle and control of P. infestans were discussed in more detail. The experiments first explained why no P. infestans isolates were found in the 2019 growing season. Then the influence of temperature on mycelium growth and spore germination on different isolates was investigated. Finally, a test was set up in which pest sensitivity was tested in different breeds.
From the temperature tests it was decided, based on all isolates together, that mycelium growth rate is maximal at 17.02 °C and 20.24 °C was the optimal temperature for spore germination. Although the reaction of the isolates relative to the different temperatures was similar, there were differences. These differences can have an influence on the occurrence of certain isolates in certain regions.
From the trial on varietal susceptibility it was clear that there are significant differences between the varieties in terms of susceptibility to lesion development on the leaves. In addition to yield and quality, this is an important factor that should be taken into account when choosing a potato variety.
1
Inhoudsopgave
Inhoudsopgave ... 1
Lijst met figuren en tabellen ... 3
Inleiding ... 5
1 Literatuurstudie ... 7
1.1 Hoofdstuk 1: Geschiedenis en belang aardappelteelt ... 7
1.1.1 Geschiedenis ... 7
1.1.2 Belang ... 8
1.1.2.1 Europa ... 9
1.1.2.2 Andere functies ... 10
1.2 Hoofdstuk 2: Systematiek en diversiteit van P. infestans populatie ... 10
1.2.1 Systematiek ... 10
1.2.2 Diversiteit van P. infestans populatie ... 11
1.2.2.1 Hoe worden de isolaten geïdentificeerd? ... 11
1.2.2.2 Verschillende genotypen in België en omringende landen ... 12
1.3 Hoofdstuk 3: Levenscyclus Phytophthora infestans ... 16
1.3.1 Levenscyclus ... 16 1.3.1.1 Geslachtelijke vermeerdering ... 20 1.3.1.2 Schadebeeld ... 20 1.4 Hoofdstuk 4: Beheersing ... 21 1.4.1 Preventie ... 23 1.4.2 Fungiciden ... 23 1.4.3 Waardplantresistentie ... 26 1.4.3.1 Inleiding ... 26 1.4.3.2 GGO ... 27
1.4.3.3 Voorbeelden van aardappelrassen met resistentie tegen P. infestans ... 28
1.4.4 Biologische organismen ... 28
1.4.4.1 Bacteriën ... 29
1.4.4.2 Schimmels... 32
1.4.5 Plantenextracten ... 33
2 MATERIAAL en METHODEN ... 35
2.1 Sampling van P. infestans isolaten ... 35
2
2.1.2 Isolaten bekomen ... 35
2.1.3 FTA-kaarten ... 36
2.2 Proef myceliumgroeisnelheden bij verschillende temperaturen ... 37
2.3 Proef sporenkieming bij verschillende temperaturen ... 37
2.4 Rasgevoeligheid ... 38
2.5 Data verwerking ... 38
3 RESULTATEN ... 40
3.1 Weeromstandigheden tijdens groeiseizoen 2019 ... 40
3.2 Effect van temperatuur op myceliumgroei ... 42
3.3 Effect van temperatuur op sporenkieming ... 45
3.4 Rasgevoeligheid ... 46
4 DISCUSSIE ... 48
5 ALGEMEEN BESLUIT ... 50
6 Literatuurlijst ... 51
3
Lijst met figuren en tabellen
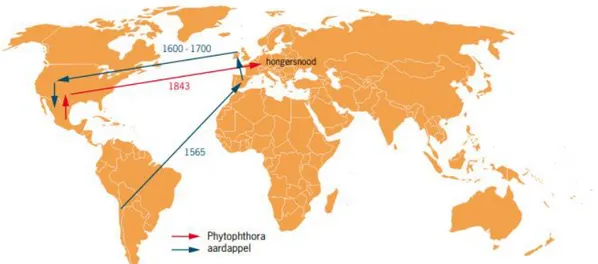
Figuur 1: Verspreiding van de aardappel en Phytophthora (Bron: Boonekamp et al., 2010). . 8
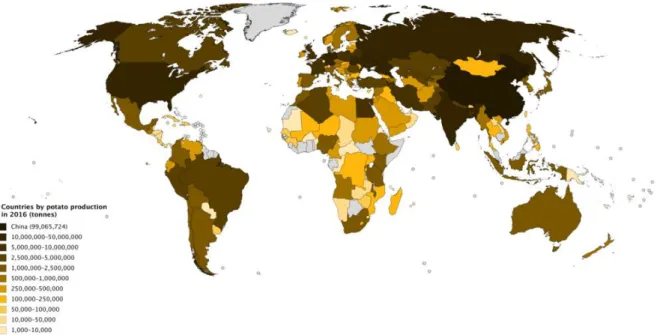
Figuur 2: Aardappelproductie per land in ton (Bron: Wikipedia). ... 9
Figuur 3: Frequentie van de verschillende P. infestans genotypen in Europa (boven) en in België (onderaan) over de jaren heen (Bron: EuroBlight website: https://agro.au.dk/forskning/internationale-platforme/euroblight/pathogen-monitoring/genotype-frequency-chart/). ...15
Figuur 4: Levenscyclus P. infestans (Bron: Drenth & Guest, 2004)...16
Figuur 5: Infectieproces P. infestans (Bron: Leesutthiphonchai et al., 2018). ...19
Figuur 6: Schadebeeld ten gevolge van P. infestans op aangetaste knollen (Bron: https://www.agro.basf.be/nl/Diensten/Ziekten-en-plagen/Aardappelziekte.html). ...20
Figuur 7: Schadebeeld ten gevolge van P. infestans aan de bladonderkant van de aardappelplant (Bron: https://wiki.groenkennisnet.nl/display/BEEL/Aardappelziekte+-+aardappel). ...21
Figuur 8: Schadebeeld ten gevolge van P. infestans aan de stengel van de aardappelplant (Bron: https://www.mooiemoestuin.nl/groenteteelt/overig/aardappel/phytophthora-infestans/). ...21
Figuur 9: Grafiek die aantoont dat de aardappelziekte jaarlijks vroeger in het veld te vinden is, seizoenen 1992-2013 (Bron: VIB dossier schimmelresistente aardappel, 2014). ...22
Figuur 10: Beschermingsindex tegen de aardappelplaag waargenomen na bladbespuiting van Pseudomonas spp. (grijs) en Bacillus spp. (wit) op het gevoelige bintje ras (Bron: Caulier et al., 2018). ...30
Figuur 11: Opbrengst bekomen na een veldproef met het challenger ras waarbij links de controle, daarna vier bacteriën en de laatste zijn twee fungiciden (Bron: Caulier et al., 2018). ...30
Figuur 12: Schimmelpluis uitgegroeid tussen twee aardappelhelften. ...36
Figuur 13: FTA kaart...37
Figuur 14: Opzet proef om effect temperatuur op sporenkieming te meten. ...38
Figuur 15: De temperatuur uitgezet in de periode mei–oktober voor groeiseizoen 2019 en 2016 (Bron: KMI). ...40
Figuur 16: De relatieve vochtigheid uitgezet in de periode mei–oktober voor groeiseizoen 2019 en 2016 (Bron: KMI). ...41
Figuur 17: De neerslag uitgezet in de periode mei–oktober voor groeiseizoen 2019 en 2016 (Bron: KMI). ...42
Figuur 18: Myceliumgroei bij isolaat Blue bij respectievelijk 4 °C, 12 °C, 20 °C en 26 °C. ...42
Figuur 19: Myceliumgroei bij isolaat EU1 bij respectievelijk 4 °C, 12 °C, 20 °C en 26 °C. ...43
Figuur 20: Myceliumgroei bij isolaat EU37 bij respectievelijk 4 °C, 12 °C, 20 °C en 26 °C. ...43
Figuur 21: Myceliumgroei bij isolaat Pink bij respectievelijk 4 °C, 12 °C, 20 °C en 26 °C. ...43
Figuur 22: Effect van verschillende P. infestans isolaten (Bleu, EU1, EU37 en Pink) op de myceliumgroeisnelhied bij temperatuur (4 °C, 12 °C, 20 °C en 26 °C). Verschillende letters wijzen op significante verschillen volgens een Tukey test. ...44
4 Figuur 23: Effect van temperatuur (4 °C, 12 °C, 20 °C en 26 °C) op de myceliumgroeisnelhied van verschillende P. infestans isolaten (Bleu, EU1, EU37 en Pink). Verschillende letters wijzen op significante verschillen volgens een Tukey test. ...44 Figuur 24: Groeicurve voor myceliumgroei (R² = 0.79), het optimum voor myceliumgroei ligt bij 17.02 °C. ...45 Figuur 25: Netto Optische densiteit (als maat voor sporenkieming) bij verschillende temperaturen (4 °C, 12 °C, 20 °C en 26 °C) voor de verschillende P. infestans isolaten (Bleu, EU1 en Pink). Verschillende letters in deel b wijzen op significante verschillen in OD tussen de temperaturen volgens een Tukey test. ...46 Figuur 26: Netto Optische densiteit (als maat voor sporenkieming) het optimum voor sporenkieming ligt op 20,24 °C. ...46 Figuur 27: Blaadjes van het ras Ratte.. ...47 Figuur 28: Spreiding van de ziekte-index (%) voor P. infestans aantasting bij de verschillende rassen. Verschillende letters wijzen op significante verschillen tussen de rassen volgens een Tukey test. ...47
Tabel 1: Genotypen van isolaten gevonden in verschillende Europese landen in 2012 Bron: Meier-runge et al., 2014). ...14 Tabel 2: Fungiciden tabel waarbij elk product gescoord wordt op verschillende parameters (Bron: https://agro.au.dk/forskning/internationale-platforme/euroblight/control-strategies/late-blight-fungicide-table/). ...25 Tabel 3: Aanbod biologische aardappelrassen in 2019 (Bron: https://www.boerderij.nl/Akkerbouw/Achtergrond/2019/2/Snelle-vooruitgang-phytophthora-resistentie-389024E/). ...28 Tabel 4: Overzicht van mogelijke alternatieven om P. infestans via biologische organismen te bestrijden. ...33 Tabel 5: Overzicht van de neerslagsom (mm) voor groeiseizoen 2019 en 2016 (Bron: KMI). ...42 Tabel 6: Netto OD voor drie isolaten (Blue, Pink en EU1) met twee mediums (Rye) bij vier temperaturen (4, 12, 20 en 26 °C). ...45 Tabel 7: Proef groeisnelheid bij verschillende temperaturen waarbij de begin diameter (Dia0), lengte diameter (Dia1a), breedte diameter (Dia1b), gemiddelde tussen breedte en lengte diameter (GemDia1) en als laatste de groeisnelheid is weergegeven. ...58
5
Inleiding
In deze masterproef werd initieel de diversiteit van P. infestans in Vlaanderen bestudeerd, maar door de voor P. infestans ongunstige weersomstandigheden kon de populatie in Vlaanderen niet in kaart gebracht worden. De alternatieve proeven stonden in het teken van waarom er geen aardappelplaag gevonden werd in de warme zomer van groeiseizoen 2019. Naast de diversiteit werd de agressiviteit van genotype Blue_13 bekeken tegenover verschillende rassen.
De aardappel is het belangrijkste niet-graan gewas in de wereld en het vierde belangrijkste voedselgewas met een jaarlijkse productie van 368 miljoen ton. Daarnaast wordt de aardappel bijna over heel de wereld geteeld behalve in gebieden waar de temperatuur het hele jaar door te hoog of te laag is (Boonekamp et al., 2010). Aardappelen kunnen ook een vitale rol spelen in het voeden van de wereld die tegen 2050 uit 9 miljard inwoners zal bestaan (Li, 2012). Moeilijkheden bij de commerciële teelt van dit gewas kan toegeschreven worden aan de gevoeligheid voor ziekten, waarvan de aardappelziekte veroorzaakt door de oömyceet P.
infestans als de belangrijkste wordt beschouwd (Axel et al., 2012). Wereldwijd hebben aardappeltelers te maken met de aardappelziekte (Boonekamp et al., 2010). Jaarlijks zorgt P.
infestans voor grote opbrengstverliezen van naar schatting tien tot vijftien procent van de
wereldwijde aardappelproductie. Voor België alleen al zouden de opbrengstverliezen neerkomen op 55 miljoen euro (VIB, z.d.). Naast opbrengstverliezen is er ook een grote belasting voor het milieu, per jaar wordt er bijna een miljoen ton werkzame stof gespoten en dit alleen in Vlaanderen die instaat voor 50-55 % van het totale Belgische aardappelareaal (Lenders & Deuninck, 2017).
Het grote probleem is dat P. infestans over de jaren heen veel agressiever is geworden, mede door de komst van het A2-mating type in de jaren 80, en dit bemoeilijkt de bestrijding (van Loon, 2019). De huidige beheersingstechniek is voornamelijk gebaseerd op de toepassing van pesticiden, een praktijk die de laatste decennia is toegenomen door de opkomst van agressievere stammen (Axel et al., 2012). Redenen hiervoor zijn dat binnen de P. infestans populatie agressievere isolaten zijn geselecteerd en doordat de diversiteit groter is geworden, omdat er verschillende recombinanten zijn gevormd tussen verschillende mating types die voorheen niet in hetzelfde gebied voorkwamen (VIB, z.d.; Leesutthiphonchai et al. 2018). Wat er bedoeld wordt met agressiever is dat de penetratieduur en ziektecyclus verkort is, er zijn ook meer generaties in eenzelfde groeiseizoen en een grotere temperatuurtolerantie (Axel et al., 2012). Om een geschikte beheersingsstrategie samen te stellen, is het belangrijk om te weten voor welke tegenstander je staat. Hier wordt inzicht in verkregen door de populatie van
P. infestans in kaart te brengen. Daarnaast is het ook belangrijk inzicht te krijgen in de
groeiomstandigheden die ideaal zijn voor P. infestans met het oog op het moment dat gevaarlijkst is voor aantasting. Er wordt een antwoord gezocht op volgende vragen: Is er een link tussen de weersomstandigheden in groeiseizoen 2019 en de verspreiding van P.
6 isolaten? Wat is het effect van temperatuur op de sporenkieming bij de verschillende isolaten? Komen de resultaten uit de proef met rasgevoeligheid overeen met de literatuur?
Deze masterproef bevat een theoretisch en een praktisch gedeelte. Het theoretische gedeelte omvat de literatuurstudie waar dieper ingegaan wordt op P. infestans en ingedeeld is in vier hoofdstukken. Het eerste hoofdstuk omvat de geschiedenis van zowel de aardappel als P.
infestans alsook het belang van de aardappelteelt in wereldomvang en specifiek Europa. Het
volgende hoofdstuk omvat de systematiek en de diversiteit van de P. infestans populatie. Hierbij wordt dieper ingegaan op de manier waarop isolaten geïdentificeerd worden en welke verschillende genotypen er aanwezig zijn in België en omringende landen. Hoofdstuk drie omvat de levenscyclus van P. infestans waar het belang van geslachtelijke vermenigvuldiging aangehaald word alsook het schadebeeld. In het vierde en laatste hoofdstuk wordt de beheersing van P. infestans besproken. Dit hoofdstuk is onderverdeeld in vijf aspecten namelijk: preventie, fungiciden, waardplantresistentie, biologische organismen en plantenextracten.
Na de literatuurstudie volgt het praktisch gedeelte van de masterproef bestaande uit materiaal en methoden, de resultaten gevolgd door een discussie met een algemeen besluit. De gedeelten materiaal en methoden en de resultaten bestaan elk uit vier delen waarbij materiaal en methoden een extra deel heeft waar de dataverwerking aangehaald wordt die gebruikt werd. Bij het eerste deel wordt uitgelegd hoe de sampling van P. infestans isolaten in zijn werk gegaan is alsook hoe het genotype van het isolaat bekomen werd. Dit werd niet aangehaald in de resultaten doordat te weinig isolaten verzameld werden door de voor P. infestans ongunstige weersomstandigheden. Om dit te staven werd in de resultaten de weersomstandigheden (temperatuur, RV en neerslag) van het plaaggevoelig groeiseizoen 2016 vergeleken met het droge groeiseizoen in 2019. In de daaropvolgende twee secties werden twee proeven uitgevoerd waarbij de sporenkieming en myceliumgroei werd bekeken bij verschillende temperaturen tussen verschillende isolaten. Als laatste werd een rasgevoeligheidsproef uitgevoerd waarbij Blue_13 werd geïnoculeerd via een DLA op verschillende aardappelrassen.
7
1 Literatuurstudie
1.1 Hoofdstuk 1: Geschiedenis en belang aardappelteelt
1.1.1 Geschiedenis
De geschiedenis van de aardappel start reeds eeuwen vóór Christus, maar er is geen kennis over de precieze oorsprong in vergelijking met andere gewassen zoals gerst en tarwe (Hawkes, 2017). Wel is geweten dat het gewas afkomstig is uit Zuid-Amerika en dat het verspreid werd door de Spaanse conquistadores in de 16e eeuw (Delcart, 2013). Vooral landen
als Bolivia, Peru en Chili op de hoogvlakten van de Andes gebruikten de aardappel als voornaamste voedingsbron voor de lokale bevolking. Er kwamen ook wilde soorten voor in Mexico en Centraal-Amerika, maar deze wilde soorten werden nooit intensief geteeld (Hawkes, 2017).
De Spaanse conquistadores veroverden de voorgenoemde gebieden in Zuid-Amerika en maakten zo kennis met de befaamde aardappel. De Spanjaarden vonden de aardappel lekker en voedzaam en brachten de aardappel richting Zuid-Spanje in 1565-1570, waar deze dan verbouwd werd in de regio van Sevilla. Op hun terugtocht uit Zuid-Amerika werd er ook opgemerkt dat de aardappel een probaat middel was tegen scheurbuik (Delcart, 2013). Door het verder telen van de aardappel werd er een voorraad opgebouwd. Deze voorraad werd dan verder verspreid naar Italiaanse kloosters die grote belangstelling toonden voor de nieuwe vrucht. Het opkomende protestantisme in ons land speelde een belangrijke rol bij de introductie van de aardappel in België. Zo stuurde het Vaticaan pater Bonomi als pauselijk legaat naar ons land om de andersgezinde gelovigen tot de orde te roepen. Pater Bonomi was ernstig ziek en kreeg aardappelen toegezonden op het adres waar hij verbleef bij Philippe de Sivry, de prefect van Mons, om zijn gezondheid te herstellen. Na het overleiden van pater Bonomi bracht Sivry het pootgoed naar de bekende botanicus Carolus Clusius die het uitvoerig bestudeerde en verder verbouwde (Delcart, 2013). Via het doorgeven van kruidkundige naar kruidkundige kwam de aardappel terecht in heel België, dit was rond 1600 (Hawkes, 2017). De aardappel werd voedingsbron nummer één bij het volk, omdat deze goed groeit in arme bodems en de aardappel kan ook direct na oogst verwerkt worden (Delcart, 2013). Door het overvloedig telen met weinig gewasrotatie werd de aardappel slachtoffer van zijn eigen succes. De eerste ernstige ziekte die voorkwam was de krulziekte die voor aanzienlijke opbrengstschade zorgde. Maar nog belangrijker, en zeker voor deze masterproef, was de Phytophthora infestans besmetting in 1845 tot 1850 die de aardappeloogst bijna geheel vernietigde, men gelooft dat tot wel 90 % van de oogst mislukte (Govers, 2010). De Ierse hongersnood in de 19de eeuw
kostte het leven van naar schatting een miljoen mensen die afhankelijk waren van de aardappeloogst. De aardappel was voor de Ieren hun belangrijkste voedselbron die tot 80 % van de nationale calorie-inname in beslag nam (Food and Agriculture Organization of the United Nations, 2008). Veel boeren en armere gemeenschappen die afhankelijk waren van de knollen waren genoodzaakt te vluchten naar de ‘nieuwe wereld’, dit was de Verenigde Staten. Niet alleen in Ierland werd de oömyceet een echte plaag, maar ook geheel West-Europa kreeg het zwaar te verduren (Govers, 2010). Figuur 1 visualiseert de verspreiding van de aardappel en van de oömyceet Phytophthora.
8
Figuur 1: Verspreiding van de aardappel en Phytophthora (Bron: Boonekamp et al., 2010).
De voorouder van de aardappelen S. Andigena die vooral in het Andes-gebergte werd verbouwd, maar zeer gevoelig was aan de aardappelplaag, werd vervangen door het bekende
S. Tuberosum afkomstig uit Chili (van Loon, 2019). Later op de tijdlijn is men gaan kijken om
meer productievere en meer ziektebestendige rassen te gaan vormen om rampen zoals de Ierse hongersnood te vermijden. Deze ziektebestendige rassen worden door veredelaars geproduceerd door nieuwe aardappelrassen uit Chili te importeren en te gaan kruisen met bestaande rassen (FAO, 2008). Het telen van een resistenter ras zal de landbouwer meer oogstzekerheid geven.
1.1.2 Belang
De aardappel is het belangrijkste niet-graan gewas in de wereld en het vierde belangrijkste voedselgewas naast rijst op nummer één, tarwe op nummer twee en maïs op nummer drie voor de mens. De aardappel wordt bijna over heel de wereld geteeld behalve in gebieden waar de temperatuur het hele jaar door te hoog of te laag is (Boonekamp et al., 2010). In 2018 was de jaarlijkse aardappelproductie 368 miljoen ton over heel de wereld bekeken. Voor rijst, tarwe en maïs was dit 782 miljoen ton, 734 miljoen ton en 1148 miljoen ton, respectievelijk. Voor tarwe is de volledige 734 miljoen ton niet voor menselijke consumptie bestemd, maar moet er 40 à 50 % afgetrokken worden die bestemd is voor veevoeder (FAO, 2020). Bij maïs is 2/3 bestemd voor veevoeder (Landing, 2018). Het internationale jaar van de aardappel werd uitgeroepen in 2008 waarbij men het belang van de aardappel voor de mensheid benadrukte. De aardappel levert voedselzekerheid en economische vooruitgang in Azië, Afrika en Latijns-Amerika, waar aardappelen een belangrijk basisvoedsel en cash crop zijn. Aardappelen kunnen ook een vitale rol spelen in het voeden van de wereld die tegen 2050 uit 9 miljard inwoners zal bestaan (Li, 2012). Op Figuur 2 wordt de wereldkaart weergegeven waarop de aardappelproductie per land in ton te zien is in 2016. Ons land behoort dus tot de landen met een zeer hoge aardappelproductie.
9
Figuur 2: Aardappelproductie per land in ton (Bron: Wikipedia).
1.1.2.1 Europa
Binnen Europa (2018) is de aardappelproductie vooral geconcentreerd in zeven lidstaten: Duitsland, Frankrijk, Polen, Nederland, Verenigd Koninkrijk, België en Roemenië. Deze zeven landen hebben ongeveer 80 % van de totale Europese aardappelproductie in handen. Nu het Verenigd Koninkrijk uit de EU is gestapt, moet er 14 % afgetrokken worden van de totale aardappelproductie (De Cicco & Jeanty, 2019). België staat op de zevende plaats met de UK erbij met 5,5 % van het Europese aardappelareaal. In de EU28 werd er in 2018 op 1,7 miljoen hectare aardappelen geteeld. Als we dit vergelijken met het totaal aantal hectaren landbouwgrond in Europa kwam dit overeen met 1,6 % van alle landbouwgrond in Europa. Dit aandeel was wel veel hoger in ons land met 11,1 % van het totale landbouwareaal beteeld met aardappelen en voor Nederland was dit 15,8 %. Volgens De Cicco & Jeanty (2019) is het aardappelareaal in de EU er wel op achteruit gegaan in de periode 2000-2018, zelfs bijna gehalveerd. Dit komt vooral door een sterke daling in Polen met 76 % en in Roemenië met 37 %. Er zijn ook landen waar het landbouwareaal voor aardappelen gestegen is namelijk in Frankrijk en in België.
In 2018 werd er in Europa 52 miljoen ton aardappelen geoogst, als dit vergeleken wordt met het jaar 2000 zitten we met een daling van 37,3 %. Desondanks de halvering van het landbouwareaal in Europa is de aardappelproductie maar gedaald met 37,3 %. Dit wil dus zeggen dat de manier waarop aardappelen worden gekweekt intensiever is geworden. Dit komt enerzijds, omdat er veel vooruitgang geboekt is in aardappelrassen en teeltmethoden. Bij de vooruitgang in aardappelrassen wordt voornamelijk gefocust op resistentie tegen ziektes zoals Phytophthora infestans, wrattenziekte, virussen, aardappelmoeheid en schurft, maar er wordt natuurlijk ook gestreefd naar een hoge opbrengst en goede kwaliteitskenmerken. In veel landen zijn dit de voorwaarden en aardappelveredelaars hebben de taak deze kenmerken te combineren. Nederland is pionier in de aardappelveredeling (van Loon, 2019). Bij de
10 vooruitgang in teeltmethoden kan er verwezen worden naar o.a. efficiëntere bemesting, plantwijzen en gewasbescherming, dit mede door de technologische vooruitgang in machines en software.
De laatste jaren worden steeds meer precisielandbouwtechnieken in de praktijk geïntroduceerd om de opbrengst per ha te verhogen en een efficiëntere input te realiseren. Een belangrijk voorbeeld daarvan is GPS-besturing. Als we spreken over GPS-besturing in de landbouw spreken we over RTK-GPS (Real Time Kinematic). De GPS maakt gebruik van satellieten in combinatie met correctiesignalen van referentiestations. Deze referentiestations zijn eigendom van de Vlaamse overheid en maken deel uit van het FLEPOS-netwerk (Flemish Positioning Service). Dit is voor iedereen gratis toegankelijk en er bestaan momenteel 45 van deze stations in Vlaanderen (Vlaamse overheid, 2020). De posities van deze referentiestations zijn exact gekend en zo worden er correcties berekend die ontvangen worden op de tractor of de machine. Door deze innovatieve uitvinding en met behulp van de correcties kan de tractor of machine tot op 3 cm nauwkeurig bestuurd worden. Door dit hulpmiddel kunnen bijna perfecte lijnen gevormd worden en worden er zo overlappen vermeden. Een klassiek RTK-GPS systeem bestaat uit een antenne voor de ontvangst van het satelliet- en correctiesignaal; een gyroscoop om de helling te corrigeren; een terminal en een geïntegreerde hydraulische stuurmodule. Bij het spuitwerk kan dit tot 10 % besparing opleveren, kostprijs van een RTK-GPS is 15.000 à 20.000 euro (Van Beek & Delanote, 2017).
Naast de technologisch vooruitgang bij machines en software, stijgt de aardappelproductiviteit door de toename aan CO2-concentratie in de atmosfeer (Hengsdijk & Verhagen, 2013).
Volgens Hengsdijk & Verhagen (2013) zal de aardappelproductiviteit meer profiteren van een toename van de atmosferische CO2-concentratie dan dat de productiviteit wordt verminderd
als gevolg van de verwachte temperatuurstijging en lagere neerslaghoeveelheid.
1.1.2.2 Andere functies
Naast aardappelen voor voeding als frieten, kroketten, puree, chips, … heeft de aardappel nog andere functionaliteiten. Behalve voor consumptie worden aardappelen ook gebruikt als veevoeder en voor industriële toepassingen voor de productie van zetmeel, vlokken en alcohol. Dit zetmeel wordt gebruikt voor productie van lijmen, textiel, papier, bouwmaterialen, etc. (VIB, z.d.).
1.2 Hoofdstuk 2: Systematiek en diversiteit van P. infestans populatie
1.2.1 Systematiek
Rijk: Chromista Klasse: Oomycota Orde: Pythiales Familie: Pythiaceae Geslacht: Phytophthora11
P. infestans lijkt uiterlijk op een schimmel, maar behoort tot de klasse van de Oomycota. Het
verschil tussen schimmels en oömyceten is dat ze o.a. een celwand hebben die cellulose bevat en geen chitine, de cellen zijn met uitzondering van de gameten diploïd en ze hebben sporen uitgerust met 2 flagellen (Haesaert, 2018; Bartnicki-Garcia & Wang, 1984)
Als we het hebben over schimmels zijn er drie mogelijke rijken namelijk de Protozoa, Chromista en de Fungi, waarbij Phytophthora infestans tot het rijk van de Chromista behoord. Dit rijk wordt door mycologen onderverdeeld in drie klassen: Oomycota, Hyphochtriomycota en de Labyrinthylomycota (Govers, 2010). Oömyceten zijn saprotrofen, pathogenen of parasieten van verschillende planten- en diersoorten zowel in het water als op het land (Bennett & Thines, 2019). Daarnaast behoort P. infestans tot de orde van de Pythiales en de familie van de Pythiaceae. Tot de familie van de Pythiaceae behoren facultatief parasitair levende schimmels die sporocyten vormen. Er bestaan twee gedaantes namelijk als saprofyt of als (zwakte) parasiet (Haesaert, 2018). Als laatste onderdeel van de systematiek behoort
P. infestans tot het geslacht van de Phytophthora. Het geslacht wordt gekenmerkt door
economisch belangrijke species die veel bestudeerd worden. Morfologisch heeft P. infestans een coenisch, hyaline en vrij vertakt mycelium waarbij de hyfen een diameter van vijf tot zeven micrometer hebben. De hyfen groeien langzaam en hebben haakse vertakkingen. Het geslacht
Phytophthora bestaat uit 313 beschreven soorten en is ontdekt door de Bary in 1876 (Ho,
2018).
1.2.2 Diversiteit van P. infestans populatie
Om een adequate bestrijdingsstrategie op te stellen is het belangrijk om de kenmerken van de pathogeenpopulatie te kennen. In deze paragraaf worden de verschillende genotypen die terug te vinden zijn in de P. infestans populatie in België en omringende landen aangehaald en op welke manier deze genotypen geïdentificeerd worden. Veel factoren beïnvloeden de populatie van P. infestans, waaronder het klimaat, de migratie, het ras dat geteeld wordt, maar ook het mating type (Brylińska et al., 2016).
1.2.2.1 Hoe worden de isolaten geïdentificeerd?
Een eerste vaak gebruikt kenmerk om P. infestans isolaten te karakteriseren is hun reactie op het fungiciden metalaxyl (Cárdenas et al., 2012). Om een beter inzicht te krijgen worden ‘merkers’ gebruikt om het genotype van een P. infestans isolaat te identificeren en dus de populatie te karakteriseren. Traditioneel gebruikte merkers zijn de allozymen glucose-6-fosfaat isomerase (Gpi) en peptidase (Pep) (Bakonyi et al., 2002). Allozymen zijn enzymen die structureel, maar niet functioneel verschillen. Allozymen werden veel gebruikt voor de ontwikkeling van DNA gebaseerde moleculaire methoden (Cooke & Lees, 2004). Voorbeelden van DNA gebaseerde merkers zijn restriction fragment length polymorphism (RFLP) fringerprint probe waarbij men informatie heeft over 25 verschillende loci (Bakonyi et al., 2002), random amplified polymorphic DNA (RAPD) en amplified fragment length polymorphism (AFLP) (Abu-El Samen et al., 2003).
De hedendaagse techniek om isolaten te identificeren gebeurt via simple sequence repeats (SSR), ook bekend als short tandem repeats (STR) (Martin et al., 2019). Dit is een zeer
12 populaire techniek die al bestaat sinds de jaren 80. Microsatellietmerkers bestaan uit motieven van één tot zes nucleotiden die meerdere malen herhaald worden en waarbij deze een kenmerkend mutatiegedrag hebben. Door het hoge mutatiecijfer die microsatellietmerkers teweegbrengen, zijn SSR’s typisch zeer polymorf. Dit wil zeggen dat verschillende individuen variantie vertonen (Guichoux et al., 2011).
Ondanks de groeiende concurrentie van nieuwe genotyperingstechnieken blijft het gebruik van deze veelzijdige en kostenefficiënte merkers toenemen, vooral gestimuleerd door de technische ontwikkelingen inzake de SSR waardoor deze vooral veel goedkoper is geworden (Guichoux et al., 2011). SSR of microsatellietmerkers worden op grote schaal gebruikt in de genetische analyse van P. infestans doordat ze een hoge reproduceerbaarheid, precisie, nauwkeurigheid en snelheid hebben en per isolaat een kleine behoefte aan genomisch DNA nodig is (Arafa et al., 2018). Microsatellieten zijn ook een beter alternatief dan de veelgebruikte dominante AFLP merkers. Dit komt doordat SSR bij nature co-dominant is waardoor deze in diploïde organismen direct onderscheidt kunnen maken tussen heterozygoten en homozygoten. Dit is belangrijk om de niveaus van inteelt af te leiden en om bepaalde allelfrequenties bloot te leggen (Dufresne et al., 2014).
Microsatellietmerkers zijn polymorfe merkers waarbij polymorf verwijst naar merkers die een zekere mate van variabiliteit in een populatie laten zien en merker verwijst naar een kenmerk dat kan worden gemeten waarbij dit in geval van microsatellietmerkers een moleculair kenmerk is (Li, 2012).
Genotypering van ziekteverwekkende populaties brengt verschillende uitdagingen met zich mee waarbij er nood is aan een gestandaardiseerde methode. Voornamelijk doordat de ziekteverwekker zich verplaatst over landsgrenzen en elk laboratorium meestal andere tests gebruikt om de populatie in kaart te brengen, waardoor de datasets niet geïntegreerd kunnen worden over de laboratoria heen. De methode moet ook snel en betaalbaar zijn als er veel isolaten moeten gegenotypeerd worden (Li et al., 2013). Li et al. (2013) heeft hierom een snelle, nauwkeurige en kosteneffectieve genotyperingmethode ontwikkeld. Hierbij wordt gebruik gemaakt van een één-staps multiplex PCR methode die gebruikt maakt van 12 microsatellietmerkers. Multiplex betekent dat er gelijktijdig meerdere microsatellietmerkers in dezelfde PCR reactie voorkomen en dit maakt de methode efficiënter (Hayden et al., 2008). Deze merkers werden geselecteerd op basis van de informatieoutput en of ze gemakkelijk te scoren zijn. Er wordt gewerkt met fluorescentie gebaseerde SSR-genotypering waarbij meerdere microsatellietmerkers tegelijkertijd in een kolom van de PCR kunnen worden gescheiden. Dit door de loci te scheiden op basis van kleuring en allelgrootte. Deze methode kan gebruikt worden om P. infestans te screenen over heel de wereld (Li et al., 2013).
1.2.2.2 Verschillende genotypen in België en omringende landen
Als men spreekt over genotypen moet er een verschil gemaakt worden tussen het A1 en A2 mating type. Voor de jaren 80 kwam alleen het A1 mating type voor in Europa. Door de introductie van het A2 mating type is geslachtelijke vermeerdering mogelijk, meer hierover in
13 sectie ‘1.3.1.1’. Algemeen zijn A2 mating types agressiever en kunnen ze oösporen vormen (van Loon, 2019).
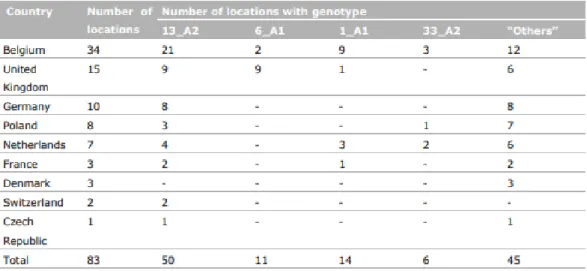
Meier-runge et al. (2014) heeft een proef opgezet in groeiseizoen 2012 waarbij er in verschillende Europese landen P. infestans stalen werden genomen. Er werd gebruik gemaakt van FTA kaarten, dit zijn chemisch behandelde filterpapieren die ontworpen zijn voor het verzamelen, conserveren en verzenden van biologische monsters voor latere DNA/RNA-analyse. De chemische behandeling zorgt voor de inactivatie van het DNA en RNA die bewaard blijven voor verdere identificatie via PCR (GD Animal Health, 2020). Per locatie werden er vier stalen genomen en de verzamelde stalen werden vervolgens gegenotypeerd via de SSR-techniek. In Tabel 1 is een overzicht weergegeven van de gevonden genotypen in 2012. In België werden er op 34 locaties stalen genomen waarvan 21 locaties het genotype 13_A2 bevatte; zes locaties het genotype 6_A1; negen locaties het genotype 1_A1; drie locaties het genotype 33_A2 en er waren 12 locaties met onbekende genotypen. Interessant is dus dat er op eenzelfde geïnfecteerde locatie meerdere P. infestans genotypen voorkomen (Meier-runge et al., 2014). Er valt ook op dat 13_A2, ook bekend als Blue_13, zeer frequent voorkomt in België en andere Europese landen.
Blue_13 is een dominante kloonlijn die gevonden werd in Europa vanaf 2004 (Li et al., 2013). In elk land van de Europese Unie werd Blue_13 gevonden vanaf 2006 (Meier-runge et al., 2014). De Blue_13-lijn behoort tot de meest agressieve lijnen op geteelde aardappelrassen, die andere agressieve lijnen in het veld overtreft en de effectieve resistentie van de plantengastheer overwint (Cooke et al., 2012). Blue_13 heeft zijn dominante positie te danken aan zijn weerstand tegen het veelgebruikte fungicide metalaxyl (Li, 2012). Genoomanalyse toonde een hoge mate van sequentiepolymorfisme en een opmerkelijk niveau van variatie in genexpressie tijdens het infectieproces, in het bijzonder van de effectorgenen. Deze polymorfismen kunnen, samen met een verlengde biotrofe fase tijdens zijn levenscyclus, de agressiviteit van Blue_13 en het vermogen om ziekte te veroorzaken op voorheen resistente aardappelrassen verklaren (Cooke et al., 2012). Uit het onderzoek van Cooke et al. (2012) bleek ook dat Blue_13 de kortste latente periode had, wat betekend dat de tijd tussen infectie en besmettelijkheid kleiner is, en dat de laesies het grootst waren. Het genotype 6_A1 bleek ook agressief.
14
Tabel 1: Genotypen van isolaten gevonden in verschillende Europese landen in 2012 Bron: Meier-runge et al., 2014).
Vaak worden de meeste locaties wereldwijd gedomineerd door één of enkele stammen, maar deze dominantie is vaak kortstondig. Studies in de Verenigde Staten en het Verenigd Koninkrijk tonen aan dat de dominante periode van een bepaalde afstamming kan variëren van één jaar tot twee tot 15 jaar (Fry, 2020).
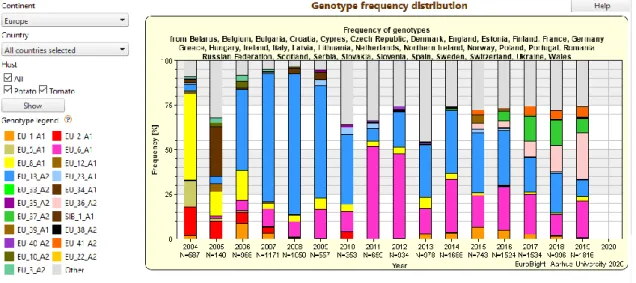
Gezien het belang van P. infestans in de aardappelsector in Europa, werd het EuroBlight netwerk opgericht. Via dit Europees netwerk wordt sedert 2004 de P. infestans populatie in kaart gebracht, om inzicht te krijgen in de populatie-opbouw en verspreiding. Figuur 3 toont de evolutie van de P. infestans populatie in Europa en in België. Zowel op basis van de resultaten in Europa als in België zien we duidelijk een toename van het EU_36_A2 (Dark Green 36) en het EU_37_A2 (Dark Green 37) genotype. Pink 6 (EU_6_A1) wordt in Europa ook frequent teruggevonden, maar werd in België de laatste jaren (2018 & 2019) niet of nauwelijks gedetecteerd.
Specifiek voor België zien we duidelijk dat van 2007 tot 2016 het genotype EU_13_A2 ook bekend als Blue_13 domineerde op de Belgische aardappelpercelen. In 2014 kwamen nieuwe genotypen EU_36_A2 en EU_37_A2 in opmars bij onze noorderburen. Vanaf 2015 kwamen deze twee genotypen overgewaaid en domineerden fel vanaf 2017-2018. EU_37_A2 is minder gevoelig voor de actieve stof fluazinam dus is het belangrijk om meerdere actieve stoffen te combineren tijdens de fungicide bespuiting (Dieleman, 2020; Schepers et al., 2019).
In Duitsland domineerde Blue_13 vanaf 2007 t.e.m. 2017, nadien nam EU_36_A2 en EU_41_A2 de dominante positie over (2019). In Duitsland is genotype EU_37_A2 ook aanwezig, maar minder dominant dan in België (EuroBlight, 2020). Genotype EU_41_A2 is een genotype dat overgewaaid is uit de Scandinavische landen en vooral aanwezig is in het noorden van Duistland. In Noorwegen werd dit genotype voor het eerst ontdekt in 2016 (Schepers et al., 2019). In België is dit genotype nog niet aanwezig (Dieleman, 2020).
Als we kijken naar Frankrijk domineerde Blue_13 van het begin van de metingen tot op heden (2006-2019). Er valt op dat de frequentie van genotype EU_1_A1 over de hele lijn (2006-2019) rond de 8 % ligt. In 2013 domineerde Pink 6 (EU_6_A1) samen met Blue_13 waarna de
15 frequentie van Pink 6 afnam. In 2016 kwam genotype EU_37_A2 fel opdagen met een frequentie van 10 % die steeg naar 35 % in 2019. EU_36_A2 vergrootte zijn frequentie van 2 % (2018) naar 11 % (2019) (EuroBlight, 2020). Volgens Schepers et al. (2019) overheersen genotypen EU_13_A2 en EU_37_A2 in het noorden en genotypen EU_13_A2 en EU_6_A1 in het westen. Genotype EU_1_A1 werd in alle Franse gebieden gedetecteerd en EU_36_A2 werd sporadisch in het noorden gevonden. Deze bron is uit 2018, EU_36_A2 zal zich waarschijnlijk meer in het noorden uitgebreid hebben.
Bij onze noordenburen domineerde Blue_13 van 2004 t.e.m. 2016, nadien nam EU_36_A2 de dominante rol over. Genotype EU_36_A2 kwam in Nederland in 2014 (2 %) op waarna zijn frequentiepercentage geleidelijk steeg tot 67 % in 2019. Genotype EU_37_A2 kwam op in 2013 (1 %) en zijn frequentie steeg t.e.m. 2017 (15 %) waarna het daalde tot 5 % in 2019 (EuroBlight, 2020).
Figuur 3: Frequentie van de verschillende P. infestans genotypen in Europa (boven) en in België (onderaan) over de jaren heen (Bron: EuroBlight website: https://agro.au.dk/forskning/internationale-
16
1.3 Hoofdstuk 3: Levenscyclus Phytophthora infestans
1.3.1 Levenscyclus
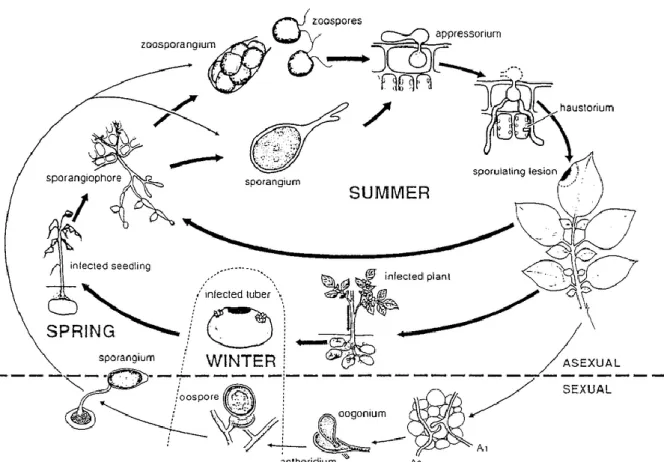
In Figuur 4 is een samenvatting weergeven van de levenscyclus van P. infestans. De oömyceet koloniseert niet alleen de aardappel en de tomaat, maar ook andere Solanaceae met name wilde gastheren zoals Solanum dulcamara, S. sarrachoides en planten van het geslacht Petunia sp. (Leesutthiphonchai et al., 2018). Bij de aardappelplant tast P. infestans zowel het blad, de stengel als de knol aan. Om te beginnen moet er een verschil gemaakt worden tussen seksuele en aseksuele vermeerdering. De seksuele vermeerdering kan alleen plaatsvinden tussen mating type A1 en A2, hier meer over in sectie “geslachtelijke vermeerdering”. In de levenscyclus van de oömyceet komen er twee aseksuele sporenvormen voor namelijk zoösporen en sporangiosporen en één seksuele sporenvorm namelijk de oösporen (Drenth & Guest, 2004).
Figuur 4: Levenscyclus P. infestans (Bron: Drenth & Guest, 2004).
P. infestans is een hemi-biotroof organisme, dit wil zeggen dat de oömyceet leeft op levende
plantencellen, maar het heeft in zijn levenscyclus ook een necrotische fase waarbij de oömyceet voedingsstoffen uit afgedode cellen haalt (Leesutthiphonchai et al., 2018). Als P.
infestans terechtkomt op een blad van een aardappelplant start het als een biotrofe schimmel,
deze fase is relatief kort. Het zal haustoria vormen in de cellen van het blad, deze haustoria dienen als zuigorganen waarmee de oömyceet voedingsstoffen zal weghalen uit de apoplast of extrahaustoriale ruimte. Na de penetratie in de cellen zal de oömyceet snel necrotroof
17 worden en zal de plant verder koloniseren door hyfen te vormen. Bij donker en vochtig weer, d.w.z. een relatieve vochtigheid (RV) van ongeveer 100 %, zullen er aan de onderkant van het blad sporangioforen vormen die door de stomata van het blad naar buiten treden (Leesutthiphonchai et al., 2018). Sporangioforen zijn de dragers van een sporangium, waarbij een sporangium instaat voor de productie van sporen. Per stomata gaan er één à vier sporangioforen naar buiten treden waaraan er sporen worden gevormd in een sporangium, terwijl de sporendrager verder groeit. De sporen in het sporangium kunnen zich ontwikkelen op twee manieren, de eerste vorm is als conidia, die direct kiemt met een kiembuis. Hiervoor is een hoge temperatuur een vereiste met een optimum bij 24 °C. Als tweede vorm kan het sporangium zich gedragen als een zoösporangium waaruit vier tot acht zoösporen met twee ongelijke flagellen tevoorschijn komen. Deze vorm komt voor bij lagere temperaturen met een optimum bij 12 °C. Deze zoösporen kunnen in een waterfilm op een blad opnieuw een infectie veroorzaken. De zoöspore zal zijn twee flagellen verliezen, krijgt een wand en zal een kiembuis vormen met een appressorium. Dit appressorium wordt gevormd aan het einde van de kiembuis. De penetratie gebeurt via huidmondjes. Door de waterdruk die het appressorium vormt, wordt de penetratiehyfe door het huidmondje geduwd. De volgende stap is het vormen van een intracellulaire infectie vescicule waaruit hyfen uitgroeien en haustoria ontstaan die voedingsstoffen gaan opnemen. Na deze stap wordt P. infestans necrotisch. De infectie is pas geslaagd als er een voldoende omvangrijk intercellulair mycelium is gevormd en als er nieuwe sporangiavorming is zodat de cyclus opnieuw kan beginnen. Gemiddeld genomen is de tijd tussen infectie en sporangiavorming zeven tot tien dagen en dit bij gunstige omstandigheden, nl. koel en vochtig weer. Door deze korte cyclus slaagt P. infestans erin om meerdere van deze cycli te doorlopen per groeiseizoen (Haesaert, 2018). Volgens Leesutthiphonchai et al. (2018) kan sporulatie al binnen de drie tot vier dagen plaatsvinden afhankelijk van het P.
infestans genotype, de waardplant en de omgevingstoestand.
Bij geslachtelijk voortplanting worden er oögonia en antheridia gevormd die respectievelijk vrouwelijke en mannelijke gameten produceren. Na de bevruchting wordt er in het oögonium een oöspore gevormd. Deze oöspore kan kiemen met een kiembuis en volgt dezelfde stappen zoals beschreven voor de zoöspore, waarbij op het einde een zoösporangium wordt gevormd (van Loon, 2019).
De ziektecyclus weergegeven in Figuur 4 kan opgesplitst worden in 12 stappen: 1. Aanhechting spore
2. Penetratie 3. Vestiging 4. Kolonisatie
5. Symptoomvorming
6. Ontwikkeling van sporendragers 7. Ontwikkeling van sporen
8. Rijping van de sporen 9. Loskomen van de sporen 10. Levensduur van de sporen
18 11. Vorming van kiemhyfe
12. Appressorium 13. Penetratie
Elke stap hierboven heeft elk zijn specifieke omgevingsvoorwaarden:
o Stadia 2-5: dit is de incubatie die in functie van de temperatuur enkele dagen kan duren. o Stadia 6-7: hiervoor is donker weer en een RV van 100 % nodig, sporenvorming
verloopt optimaal bij 16-22 °C. o Stadium 8: hier is licht gewenst.
o Stadium 9: sporen komen los door een daling in RV doordat er torsie optreedt bij de sporendragers.
o Stadium 10: de levensduur van de sporen is afhankelijk van de weersomstandigheden, bij gematigde weersomstandigheden kunnen sporen enkele kilometers vitaal blijven. o De tijd tussen de kieming en infectie is temperatuur afhankelijk, nl. bij 10-15 °C duurt
de kieming 0,5-2 u; bij 15-25 °C duurt het 2-2,5 u en boven de 33 °C is er geen groei meer. Dit alles kan pas plaatsvinden als er een waterfilm op het blad aanwezig is. De hierboven vermelde data zijn gemiddelden daar elk genotype en zelfs elk isolaat zijn eigen kenmerkende epidemiologische karakteristieken heeft (Haesaert, 2018; CABI, 2020)
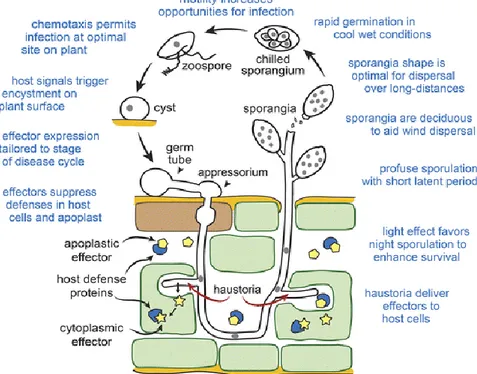
Figuur 5 geeft de ziektecyclus in meer detail weer van P. infestans vanaf het moment dat de pathogeen een blad infecteert en toont tevens aan waarom P. infestans zo’n succesvolle pathogeen is. De meeste infecties in een groeiseizoen gebeuren door de aseksuele sporangia daarom wordt er hier dieper op in gegaan. Zoals voorheen vermeld gaan de sporangia aan de onderkant van het blad door een huidmondje groeien. Deze sporangia kunnen door de wind in 3 uur tijd 10 tot 20 km meegevoerd worden (Leesutthiphonchai et al., 2018). Volgens Leesutthiphonchai et al. (2018) bestaat er een chemotaxis tussen het bladoppervlak van een aardappelplant en de sporen afkomstig van P. infestans. Hierdoor worden de sporen aangetrokken door wondjes op het bladoppervlak waardoor de infectie optimaal kan plaatsvinden.
Eenmaal in de plant gaat P. infestans zich uitbreiden en gaat het vooral om intercellulaire hyphae die haustoria in de gastheercellen vormt. Bij het nestelen zal de plant dit herkennen en zich ertegen verzetten, maar P. infestans zal hier natuurlijk gepast op reageren. Zoals vele dierlijke en plantaardige ziekteverwekkers gaat P. infestans eiwitten afscheiden, zogenaamde effectoren genoemd (Figuur 5), die de kolonisatie van de oömyceet vergemakkelijken. Deze effectoren gaan de fysiologie van de gastheerplant veranderen en gaan de immuniteit onderdrukken. De parasitaire effectoreiwitten richten zich op verschillende plaatsen in het plantenweefsel van de gastheerplant. Er bestaan apoplastische effectoren welke in de extracellulaire ruimte handelen en interfereren met de apoplastische plantenverdediging zoals proteasen en glucanasen. Ten tweede wordt er een groot aantal cytoplasmatische effectoren in de gastheercellen gepompt door P. infestans die de gastheereiwitten in het cytoplasma van de cel gaat proberen inhiberen (Raffaele et al., 2010). Figuur 5 geeft een voorstelling van de twee verschillende effectoren en hoe deze de gastheereiwitten onderdrukken. P. infestans is
19 ook instaat om de verdediging van de gastheer te ontwijken, dit omdat de celwand van de schimmel geen chitine bevat en planten zoeken naar chitine om schimmels op te sporen. Een tweede manier die P. infestans gebruikt om de verdediging te slim af te zijn, is door de schade aan de gastheercellen te beperken tijdens zijn biotrofe fase en zo dus onopgemerkt blijft voor het afweersysteem van de plant. Dit doet de oömyceet door alleen voedingsstoffen uit de apoplast en extrahaustoriale membraan te halen (Leesutthiphonchai et al., 2018).
Sporangia productie gebeurt niet bij toeval als het donker is en aan de onderkant van het bladoppervlak. De sporangia doen dit om de overlevingskansen te vergroten. Doordat sporangia gehydrateerd en hyalien (glasachtig doorzichtig) zijn, zijn ze gevoelig voor uitdroging en UV-beschadiging (Leesutthiphonchai et al., 2018). De zoösporen hebben ook vocht nodig om vrij te komen uit het sporangium, daarom is dauw het ideale moment om te ontluiken. Xiang & Judelson (2014) hebben proeven uitgevoerd waarbij ze verschillende licht regimes hanteerden bij constante temperatuur en constante relatieve luchtvochtigheid. Er werd geconcludeerd dat P. infestans zijn sporulatiepatroon aanpaste aan dat van de veranderde licht regimes en als er geen donkere periode is, dan zal de sporulatie toch doorgaan. P. infestans bevat lichtgevoelige transcriptiefactoren die het sporulatiepatroon regelen. Meestal bevatten fungi UV-blokkerende pigmenten, zoals carotenen, maar P.
infestans heeft deze pigmenten niet (Leesutthiphonchai et al., 2018).
Figuur 5: Infectieproces P. infestans (Bron: Leesutthiphonchai et al., 2018).
De overwintering van P. infestans kan op twee manieren gebeuren. De eerste manier ontstaat als er alleen ongeslachtelijk vermeerdering kan plaatsvinden. In dit geval gebeurt de overwintering via besmette knollen die na de oogst achterbleven op de akker of via besmette knollen aanwezig in het pootgoed en afvalhopen. Een tweede manier is de overwintering als oöspore die ontstond via geslachtelijke vermeerdering. Deze oösporen kunnen twee tot drie jaar vitaal blijven in de bodem (Haesaert, 2018). Oösporen zijn veel persistenter dan
20 ongeslachtelijke sporen, oösporen kunnen vriestemperaturen en fungiciden overleven (Leesutthiphonchai et al., 2018).
1.3.1.1 Geslachtelijke vermeerdering
Een nieuwe reeks problemen ontstond in de jaren 80 wanneer er voor het eerst in Europa het A2-mating type van P. infestans werd gevonden. Volgens van Loon (2019) is de meest waarschijnlijke oorzaak van hoe dit is kunnen gebeuren, de import van aardappelen in het voorjaar van 1977. Deze import was nodig doordat er in Europa een tekort was als gevolg van droogte in het jaar 1976. Het A2-mating type kwam voorheen niet voor in Europa, maar door de introductie kan P. infestans nu dikwandige oösporen vormen. Deze oösporen zijn dus een eerder recent verschijnsel en zorgen er voor dat P. infestans één tot drie jaar in de bodem kan overleven. Daarom is in België een minimale rotatie van één op drie verplicht voor aardappelen. Door een te enge vruchtwisseling zou er een massale verspreiding van de oömyceet ontstaan. Het A2-mating type heeft P. infestans dus nodig om zich seksueel voor te kunnen planten (van Loon, 2019). Seksuele voorplanting resulteert ook in meer genetische variatie waardoor dit gaat leiden tot een verhoogde en snellere evolutie van P. infestans. Het genotype Blue_13 is hier een mooi voorbeeld van, het bouwde snel resistentie op tegen het veelgebruikte fungicide metalaxyl en domineerde de Europese aardappelpercelen vanaf 2006 (Li, 2012) (EuroBlight, 2020). Door de seksuele voorplanting hebben meer agressieve stammen zich ontwikkeld met kortere levenscycli (Axel et al., 2012).
1.3.1.2 Schadebeeld
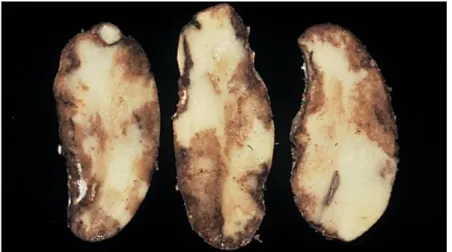
Het zeer epidemisch karakter van P. infestans is deels te wijten aan het vermogen om alle organen van de plant (bladeren, stengels en knollen) aan te tasten, in alle stadia van de gewasontwikkeling zolang de klimatologische omstandigheden gunstig zijn (Gaucher et al., 2018). Figuur 6 toont het schadebeeld op aangetaste knollen. Onder deze vorm kan P.
infestans de winter overleven.
Figuur 6: Schadebeeld ten gevolge van P. infestans op aangetaste knollen (Bron: https://www.agro.basf.be/nl/Diensten/Ziekten-en-plagen/Aardappelziekte.html).
Figuur 7 illustreerd de laesies (bladvekken) waarop het ‘wit pluis’ te zien is aan de bladonderzijde van de aardappelplant. Dit wil zeggen dat de oömyceet nog actief is en komt enkel voor als de luchtvochtigheid hoog genoeg is. Het schimmelpluis bestaat uit
21 sporendragers en de daarop gevormde sporen. Normaal komt dit schimmelpluis alleen voor aan de bladonderzijde, maar er bestaan ook stammen die aan beide zijden kunnen sporuleren. In een vergevorderd stadium kan P. infestans het gehele blad inpalmen (PCA, 2012).
Figuur 7: Schadebeeld ten gevolge van P. infestans aan de bladonderkant van de aardappelplant (Bron: https://wiki.groenkennisnet.nl/display/BEEL/Aardappelziekte+-+aardappel).
Op de stengel van de aardappelplant kunnen grote langwerpige grauwbruine tot bruinzwarte laesies ontstaan, die in een vergevorderd stadium de gehele stengel kan omvatten (Figuur 8). Als de luchtvochtigheid hoog genoeg is, ontstaat er, net als bij het blad, wit schimmelpluis. Het enigste verschil is dat aangetast stengelweefsel meermaals kan sporuleren. Stengellaesies ontstaan voornamelijk uit bladoksels, omdat deze moeilijk opdrogen (PCA, 2012).
Figuur 8: Schadebeeld ten gevolge van P. infestans aan de stengel van de aardappelplant (Bron: https://www.mooiemoestuin.nl/groenteteelt/overig/aardappel/phytophthora-infestans/).
1.4 Hoofdstuk 4: Beheersing
Jaarlijks zorgt P. infestans voor grote opbrengstverliezen van naar schatting tien tot vijftien procent van de wereldwijde aardappelproductie. Voor België alleen al zouden de opbrengstverliezen neerkomen op 55 miljoen euro (VIB, z.d.). Naast opbrengstverliezen is er ook een grote belasting voor het milieu. Volgens het VIB, Vlaams Instituut voor Biotechnologie, wordt er in Vlaanderen gemiddeld 17 kg werkzame stof per hectare per jaar gespoten om P.
22
infestans te bestrijden. Dit is natuurlijk zeer afhankelijk van het groeiseizoen. In groeiseizoen
2018 en 2019 was er amper aardappelplaag en zal er dus veel minder werkzame stof gebruikt zijn. Volgens het departement Landbouw en Visserij bedroeg het aardappelareaal in 2019 in Vlaanderen 55.000 ha. Als we dit vermenigvuldigen met 17 kg wordt er per jaar bijna een miljoen ton werkzame stof gespoten en dit alleen in Vlaanderen die instaat voor 50-55 % van het totale Belgische aardappelareaal (Lenders & Deuninck, 2017).
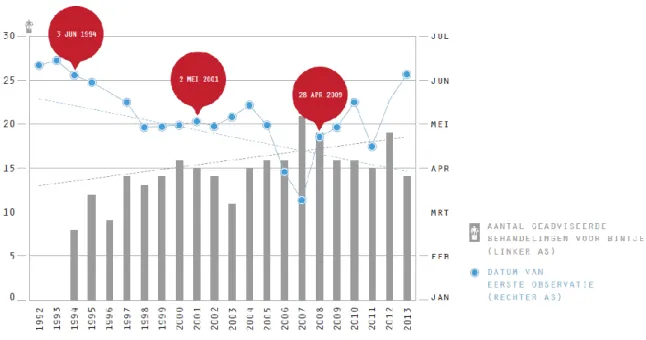
Het grote probleem is dat P. infestans over de jaren heen veel agressiever is geworden en dit bemoeilijkt de bestrijding. Redenen hiervoor zijn dat binnen de P. infestans populatie agressievere isolaten zijn geselecteerd en doordat de diversiteit groter is geworden, omdat er verschillende recombinanten zijn gevormd tussen verschillende mating types die voorheen niet in hetzelfde gebied voorkwamen (VIB, z.d.). Wat er bedoeld wordt met agressiever is dat de penetratieduur en ziektecyclus verkort is, er zijn ook meer generaties in eenzelfde groeiseizoen en een grotere temperatuurtolerantie. Optimale temperaturen zijn 10-25 °C voor de oömyceet. Figuur 9 illustreert de toegenomen agressiviteit van P. infestans waarbij de oömyceet vroeger in het groeiseizoen tevoorschijn komt. Dit zorgt natuurlijk voor een toename in het aantal bespuitingen die moeten gebeuren per seizoen.
Figuur 9: Grafiek die aantoont dat de aardappelziekte jaarlijks vroeger in het veld te vinden is, seizoenen 1992-2013 (Bron: VIB dossier schimmelresistente aardappel, 2014).
P. infestans is een dynamische ziekteverwekker die telkens opnieuw ingekruiste resistentie
omzeilt (Govers, 2010). De enorme dynamiek van P. infestans vindt zijn oorsprong in het DNA.
P. infestans beschikt over grote, flexibele gen families die zorgen voor een snelle aanpassing
aan nieuwe omstandigheden. Als er inzicht is in deze dynamiek kunnen er betere en adequatere bestrijdingsstrategieën ontwikkelt worden. Het zeer epidemisch karakter is ook te wijten aan het vermogen om alle organen van de plant (bladeren, stengels en knollen) aan te
23 tasten, in alle stadia van de gewasontwikkeling zolang de klimatologische omstandigheden gunstig zijn (Gaucher et al., 2018).
Ondanks het feit dat veel onderzoekers over gans de wereld zich concentreren op het vinden van een efficiënte en goedkope manier om P. infestans te beheersen, blijven fungiciden de belangrijkste bestrijdingsstrategie. Deze strategie is niet alleen een grote economische investering voor de landbouwer, maar is ook schadelijk voor het milieu (Cárdenas et al., 2012). Dit doordat men meestal 10-14 keer moet spuiten tegen P. infestans in een groeiseizoen.
1.4.1 Preventie
Dit is één van de belangrijkste maatregeling die de landbouwer kan nemen om P. infestans te beheersen. Door de trend van alsmaar meer agressievere stammen zal preventie in de komende jaren nog belangrijker worden, mede doordat de agressievere stammen een hogere sporendichtheid hebben, kortere levenscycli, kortere en efficiëntere incubatietijd en grotere vlekken hebben (Dieleman, 2020).
Om de ziektedruk te verlagen is het belangrijk om hygiënische maatregelen te nemen d.w.z. primaire infectiehaarden vermijden, i.e. opslagplanten bestrijden en aardappelen in afvalhopen vernietigen (VIB, z.d.). Na de oogst moeten gedeeltelijk aangetaste bladeren, knollen en stengels verwijderd worden of vernietigd worden door herbiciden om te voorkomen dat deze besmet worden met P. infestans (Goutam et al., 2018). Volgens Goutam et al. (2018) zou organisch materiaal bij het ontbinden, ammoniak en vluchtige organische zuren produceren die P. infestans zou afdoden. Stalmest uitrijden na oogsten is een optie, maar dit kan ten laatste tot 31 oktober. Vanaf 16 januari t.e.m. 31 augustus kan er bemest worden waarbij dit bij derogatiepercelen vanaf 16 februari kan. Vanaf 1 September t.e.m. 31 oktober kan het op niet-derogatiepercelen met een maximale dosis van 50 kg werkzame N/kg (VLM, 2020). Bij het opkomen van het bladerdek kan er een preventieve bespuiting plaatsvinden waarbij het belangrijk is dat alle planten geraakt zijn door het toepassen van een goede spuittechniek (Dieleman, 2020).
De oösporen, dus de geslachtelijke sporen, kunnen niet of toch niet economisch rendabel vernietigd worden. De beste manier om geslachtelijke sporen te slim af te zijn, is door een ruime vruchtwisseling omdat ze tot drie jaar vitaal blijven. Door de vruchtwisseling wordt de periode tussen de aardappelcycli verlengd. Door deze maatregelen wordt de verspreiding van
P. infestans verminderd (VIB, z.d.). Rassenkeuze is natuurlijk ook een preventieve maatregel,
meer hierover in sectie “waardplantresistentie”. Bij de rassenkeuze is het gebruik en keuze van gezonde knollen zeer belangrijk (Axel et al., 2012).
1.4.2 Fungiciden
Fungicidengebruik in de aardappelteelt neemt 10 à 20 % van de totale kosten in voor productie en is tot op heden de ‘goedkoopste’ bestrijdingstechniek (Leesutthiphonchai et al., 2018). Door het gebruik van scouting of voorspelling kan het fungicidengebruik optimaal benut worden (Axel et al., 2012). Het PCA, het Proefcentrum voor de Aardappelteelt, heeft een waarschuwingssysteem die telers alarmeert wanneer er best gespoten wordt. Volgens het PCA stijgt het aantal bespuitingadviezen en vervroegt de eerste vaststelling van de
24 aardappelplaag. Het is belangrijk dat alle werkzame stoffen beschermd worden en dat er aan resistentiemanagement wordt gedaan (Dieleman, 2020). Een lijst met alle toegelaten gewasbeschermingsmiddelen is te vinden op de site van Fytoweb.
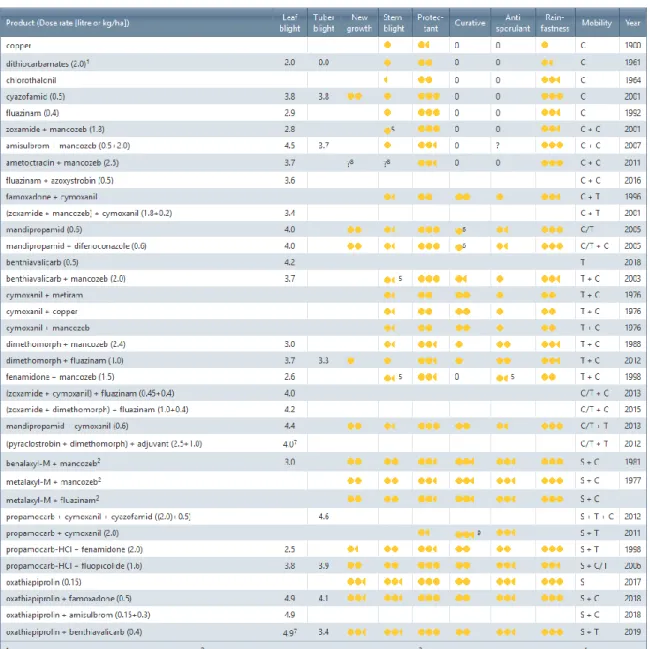
Binnen het EuroBlight netwerk werd een effectiviteitstabel ontwikkeld (Tabel 2), die elk jaar geüpdatet wordt, voor de bestrijding van P. infestans. De scores voor de bestrijding van P.
infestans zijn gebaseerd op onafhankelijke proeven uitgevoerd in meerdere jaren op
verschillende Europese locaties. De EuroBlight tabel is opgesteld om een aantal kenmerken van het product te evalueren en om een overzicht te geven over de waarde van het kenmerk. Deze tabel kan door aardappeltelers in Europa gebruikt worden om advies en ondersteuning te verkrijgen bij de besluitvorming om voor een bepaalde bestrijding te kiezen (WUR, z.d.;
EuroBlight, 2020).
De scores worden vastgesteld door experts van EuroBlight (onafhankelijke onderzoekers en afgevaardigden van de gewasbeschermingsindustrie) op basis van beschikbare gegevens. Kolom twee geeft de doeltreffendheid weer van de fungicide om bladziekte veroorzaakt door
P. infestans te bestrijden waarbij een schaal van 2-5 gehanteerd wordt. De volgende kolom is
hetzelfde, maar dan voor knolziekte waarbij een schaal van 0-5 gehanteerd wordt. Kolommen 4 t.e.m. 9 hanteren een schaal waarbij 0 geen effect voorstelt; = redelijk effect ; = goed
effect ; = zeer goed effect en Blanco wil zeggen dat er geen beoordeling heeft
plaatsgevonden. Kolom vier geeft weer in hoeverre nieuwgevormd loof beschermd wordt. De volgende kolom geeft de stengelbescherming weer. Kolom 6 t.e.m. 8 geven drie werkingsmechanismes weer. Bij de preventieve werkingswijzen worden de sporen gedood voor of tijdens de ontkieming/penetratie. Hierbij moet het fungicide aanwezig zijn op het blad/stengel voordat de sporen ontkiemen en binnendringen. Bij een curatieve werkingswijze
is het fungicide actief tegen P. infestans in de periode direct na besmetting, maar voordat de
symptomen zichtbaar worden. Bij het laatste werkingsmechanismen gaat het fungicide de P.
infestans-laesies beïnvloeden door de sporenvorming te verminderen en/of de
levensvatbaarheid van de gevormde sporen te verminderen. Regenvastheid: een fungicide wordt als regenvast beschouwd als het na gebruik voldoende is opgedroogd of opgenomen is door de plant, zodat het nog werkt na regen of beregening. Bij de voorlaatste kolom gaat het over de mobiliteit in de plant waarbij er onderscheid word gemaakt tussen contact, translaminair of systemisch. De laatste kolom stelt het introductiejaar voor van het fungicide
(WUR, z.d.; EuroBlight, 2020). Niet meer toegelaten in België vanaf 1 oktober 2019:
25
Tabel 2: Fungiciden tabel waarbij elk product gescoord wordt op verschillende parameters (Bron: https://agro.au.dk/forskning/internationale-platforme/euroblight/control-strategies/late-blight-fungicide-table/).
Er wordt meestal, om niet te zeggen altijd, een combinatie gemaakt van werkzame stoffen met een verschillende werkingswijze, waarbij vaak Mancozeb gebruikt wordt in combinatie met een nieuwere werkzame stof. Mancozeb is een werkzame stof die al sinds 1961 geregistreerd is als fungicide, die nog altijd volop gebruikt wordt en een belangrijke positie heeft in de aardappelteelt (van Doorne et al., 2018). Op dit moment is mancozeb in België toegelaten voor tal van teelten namelijk: aardappelen, tarwe, uien, peren, appels, druiven, bonen, … (Fytoweb, 2020). Mancozeb is een multi-site fungicide die niet resistentiegevoelig is en ook plantenvoedingstoffen als mangaan bevat. In de aardappelteelt wordt mancozeb veelvuldig gebruikt in de preventieve bestrijding van P. infestans en Alternaria (solani) (van Doorne et al., 2018).
26 Koper wordt gebruikt onder de vorm van koperhydroxide, koperoxychloride, kopersulfaat en koperoxide en wordt al vele jaren gebruikt. Koper is het enige chemische middel dat toegestaan is in de biologische landbouw. Koper kan op lange termijn gevolgen hebben voor het milieu vanwege de accumulatie in de bodem. Hierdoor heeft de Europese Commissie het gebruik van koper beperkt tot 6 kg koper per ha per jaar (Shtienberg et al., 2010).
1.4.3 Waardplantresistentie
1.4.3.1 Inleiding
Bij de introductie van de oömyceet P. infestans in Europa bleek dat de aardappelziekte, bij massale uitbraak, grote gevolgen had voor de teelt. Niet alleen voor de opbrengst, maar ook de visie van wat een goed ras is, werd helemaal veranderd. De veredeling was van toen af aan opzoek naar rassen die een verminderde gevoeligheid vertonen tegen de aardappelplaag. Het was niet eenvoudig om een ras te creëren die resistent was tegen P. infestans daar alle bestande rassen vooral vatbaar waren (van Loon, 2019).
Tot op heden worden er in de gangbare landbouw weinig resistente rassen tegen P. infestans op grote schaal geteeld, omdat deze rassen goede eigenschappen kwijt zijn gespeeld tijdens te veredeling. Sarpo Mira wordt wel door de industrie gebruikt. De nadelen op gebied van opbrengst en verwekingseigenschappen zijn niet rendabel genoeg om deze resistente gewassen te telen. De verwerkingseigenschappen hierboven vermeld, zijn bakkwaliteit en bewaarbaarheid. Deze zijn belangrijk voor de frietindustrie en er bestaan nog geen resistente rassen die op vlak van kwaliteit kunnen concurreren met de gangbare rassen. Ook zijn er nog niet voldoende rassen voor alle doeleinden. Voor tafelaardappelen bijvoorbeeld is er al een vrij ruim pakket aan rassen, maar deze rassen zijn niet jaarrond beschikbaar. In de biologische landbouw worden deze resistentere rassen natuurlijk wel gebruikt omdat zij geen fungiciden mogen spuiten en dus geen keus hebben (VIB, z.d.).
Waardplantresistentie is gebaseerd op herkenningsmechanismen. Men is opzoek naar resistentiegenen (R-genen) die instaat zijn resistentie te bewerkstelligen via herkenning van (specifieke) eiwitten van de ziekteverwekker, effectoren genoemd. Bij de veredeling wil men zoveel mogelijk R-genen combineren om een zo hoog mogelijke resistentie te bekomen (Vossen et al., 2007). Het verkrijgen van een breed spectrum resistentie door gebruik te maken van meerdere R-genen in aardappelgenotypen door traditionele veredeling verloopt zeer traag door het heterozygoot en tetraploïd karakter van commerciële aardappelgenotypen (Haesaert et al., 2015). Mede door de snellere generatiewissels naar nieuwe virulente stammen is er nood aan nieuwe technieken, dit omdat de klassieke veredeling te veel tijd in beslag neemt (Goutam et al., 2018).
Een andere techniek is het gebruik van ‘Systemic acquired resistance’ (SAR), hiermee kan men de immuniteit van de plant verhogen. Het verdedigingsmechanisme wordt gestimuleerd door SAR-inducers die een primaire infectie veroorzaken door ziekteverwekkers en zo het immuunsysteem boosten in opvolgende infecties (Goutam et al., 2018).
RNA silencing (RNAsi), ook bekend als RNA interferentie (RNAi), is een techniek die de laatste jaren meer aandacht krijgt in de strijd tegen de oömyceet P. infestans. RNAsi is een krachtig