FUNCTIONELE
TUINTJES
ALS
MEETINSTRUMENT
VOOR
ECOSYSTEEMDIENSTEN LANGS EEN
LANDSCHAPPELIJKE
COMPLEXITEITSGRADIËNT OP HET
PLATTELAND
Aantal woorden: 12738
Benedikte Van Heghe
Studentennummer: 01610365Promotor(en): Prof. Ir. Jan Mertens
Tutor: Ir. Frederik Gerits
Masterproef voorgelegd voor het behalen van de graad master in de biowetenschappen: land- en tuinbouwkunde (tuinbouwkunde)
i
AUTEURSRECHTELIJKE BESCHERMING
“De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te stellen en delen van de scriptie te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze scriptie.”
“The author and the promoter give the permission to use this thesis for consultation and to copy parts of it for personal use. Every other use is subject to the copyright laws, more specifically the source must be extensively specified when using the results from this thesis”.
ii
WOORD VOORAF
Eerst en vooral wil ik Prof. Ir. Jan Mertens en Ir. Frederik Gerits bedanken voor de goede begeleiding bij deze masterproef. Ze hebben mij steeds gemotiveerd, mijn vragen beantwoord en hebben dit onderzoek in goede banen geleid.
Ik ben blij dat ik het voorrecht heb gehad om mee te mogen werken aan het BEL-landschap project. Het leek me vanaf het begin een zeer interessant onderzoek en was meteen verkocht. In het bijzonder wil ik alle vrijwilligers bedanken voor de tijd die ze hebben geïnvesteerd in het onderhoud van de tuintjes en de dataverzameling. Hun bijdrage vormde het geraamte voor dit onderzoek. De resultaten van dit onderzoek zijn rechtstreeks verbonden met de zorg die ze in de tuintjes hebben gestoken. Ik wil ook de sponsors en instellingen bedanken die dit project mede mogelijk gemaakt hebben.
Verder wil ik mijn ouders bedanken voor de kansen die ze mij hebben gegeven. Ze hebben mij steeds gesteund en geholpen waar mogelijk. Ook Rafael wil ik bedanken voor de steun en het nalezen van het eindresultaat van deze thesis.
iii
SAMENVATTING
Kernwoorden: natuurlijke plaagbestrijding, bodemvallen, landschapssamenstelling, loopkevers, kortschildkevers, duizendpoten, miljoenpoten, pissebedden, mieren, spinnen
Abstract (NL)
Natuurlijke plaagbestrijding is een belangrijke ecosysteemdienst voor de landbouwsector. Deze dienst wordt geleverd door natuurlijke vijanden die afhankelijk zijn van de kwaliteit van het omringende landschap. In deze thesis wordt het effect onderzocht van het landschap en het microklimaat op grond-kruipende arthropoda. Hiertoe wordt gebruik gemaakt van een innovatieve meettechniek die steunt op burgerwetenschap. Er werden 40 gestandaardiseerde vierkante meter ‘meettuintjes’ geplaatst op locaties met verschillende landschapssamenstellingen. Deze tuintjes werden onderhouden en geoogst door vrijwilligers. De grond-kruipende arthropoda werden verzameld door in ieder tuintje een bodemval te plaatsen. De vallen werden na twee weken opgehaald, deze procedure werd drie keer herhaald (in juni, juli en augustus). Met deze methode werden niet alleen natuurlijke vijanden (loopkevers, kortschildkevers, duizendpoten en spinnen) gevangen maar ook detritivoren (pissebedden en miljoenpoten) en mieren. De landschapssamenstelling werd onderzocht a.d.h.v. het percentage landbouw, hoog groen, laag groen en niet groen binnen concentrische cirkels van 10, 50 en 500 meter. Het effect van het microklimaat werd onderzocht aan de hand van de temperatuur in de meettuintjes. Er werd gevonden dat het landschap een belangrijke invloed heeft op de samenstelling van de arthropodagemeenschap en op predatoren in het bijzonder. Onze resultaten suggereren dat niet alle natuurlijke plaagbestrijders op gelijke wijze beïnvloed worden door het omliggende landschap. De activiteit-densiteit van de eerste groep natuurlijke vijanden (loopkevers, kortschildkevers en duizendpoten) was het hoogst wanneer er veel landbouw in de omgeving van het tuintje aanwezig was. De activiteit-densiteit van de andere natuurlijke vijanden (spinnen) werd voornamelijk bepaald door afwezigheid van niet groen. De detritivoren (pissebedden en miljoenpoten) reageerden verschillend op de landschapssamenstelling. Bij pissebedden kon niet worden aangetoond dat de landschapssamenstelling de activiteit-densiteit in belangrijke mate beïnvloed. Miljoenpoten werden voornamelijk teruggevonden in tuintjes met veel hoog groen. De activiteit-densiteit van mieren werd positief beïnvloed door het percentage niet groen, dit is mogelijks door een intermediair effect van het microklimaat.
Abstract (EN)
Biological pest control is an ecosystem service from which agriculture benefits substantially. This service is brought by natural enemies who depend on the quality of the surrounding landscape. This thesis examines the effect that landscape and microclimate could have on ground-dwelling arthropods. To this end, an innovative measuring technique has been used based on ‘citizen science’. Forty standardized small ‘measurement gardens’, plots of 1m², had been placed on locations with different landscape compositions. Gardening and harvesting were done by volunteers. The ground-dwelling arthropods have been collected by placing pitfall traps in each garden. The traps were collected every two weeks. This procedure was repeated three times (throughout June, July and August). Not only natural enemies (carabid beetles, rove beetles, centipedes and spiders) were caught using this method, but also detritivores (isopods and millipedes) and ants. The landscape composition has been examined using the percentage of agriculture, high vegetation (higher than 3m), low vegetation (lower than 3m) and non-vegetation (urban land, water, barren ground), within concentric circles of 10, 50 and 500m. The effect of the microclimate has been examined measuring plot temperatures. We found an important effect of the surrounding landscape on the composition of the arthropod community and predators in particular. Our results suggest not all natural pest control agents are influenced in a similar way by the surrounding landscape. The activity density of the first group of natural enemies (carabid beetles, rove beetles and centipedes) was high if the plot was situated in an area with much agriculture. The activity density of the other natural enemies (spiders) was mainly determined by the
iv
absence of non-vegetation. Detritivores (isopods and millipedes) react differently on the landscape composition. The landscape composition could not be determined as a prominent influence on the activity density of isopods. Millipedes were mostly found in a high-vegetation area. The activity density of ants has been positively influenced by the share of non-vegetation, though this might be due to an intermediary effect of the microclimate.
v
INHOUDSOPGAVE
Auteursrechtelijke bescherming ... i Woord vooraf ... ii Samenvatting ... iiiAbstract (NL) ...iii
Abstract (EN) ...iii
Inhoudsopgave ... v
1. Lijst met tabellen en figuren ... 1
Lijst met tabellen ... 1
Lijst met figuren ... 1
2. Inleiding ... 3
3. Literatuurstudie ... 4
Ecosysteemdiensten ... 4
3.1.1.
Definitie en indeling van ecosysteemdiensten ... 4
3.1.2.
Ecosysteemdiensten en biodiversiteit ... 5
Arthropoda als leveranciers van ecosysteemdiensten: natuurlijke vijanden ... 6
3.2.1.
Coleoptera ... 6
3.2.2.
Araneae ... 7
3.2.3.
Hymenoptera ... 7
Invloed van het landschap op natuurlijke vijanden ... 8
3.3.1.
Belang van het landschap voor natuurlijke vijanden ... 8
3.3.2.
Habitatfragmentatie ... 8
3.3.3.
Habitatverlies en vermindering van de habitatkwaliteit... 9
4. Materiaal en methoden ... 10
Dataverzameling van de arthropoda en het microklimaat via functionele meettuintjes ... 10
4.1.1.
Meetinstrument: functionele tuintjes ... 10
4.1.2.
Dataverzameling van de arthropoda ... 10
4.1.3.
Dataverzameling van het microklimaat ... 10
Landschapssamenstelling 10, 50 en 500 meter rondom de tuintjes ... 11
Dataverwerking van de resultaten ... 12
4.3.1.
Transformatie van de arthropoda activiteit-densiteit ... 12
4.3.2.
PCA-analyse ... 13
4.3.3.
Opdeling in absence en presence data bij duizendpoten en miljoenpoten ... 14
vi
4.3.5.
Exploratieve data-analyse bij de data van het microklimaat ... 14
5. Resultaten ... 15
Invloed van de landschapssamenstelling op in de grond kruipende arthropoda ... 15
5.1.1.
PCA-analyse ... 15
5.1.2.
Absence-presence ... 16
5.1.3.
Meervoudige lineaire regressie ... 17
Microklimaat ... 18
5.2.1.
Verband tussen het microklimaat en de landschapssamenstelling ... 18
5.2.2.
Verband tussen arthropoda en microklimaat ... 19
6. Discussie ... 20
Wat is het effect van het percentage landbouw/niet groen/laag groen/hoog groen op
arthropoda? ... 20
6.1.1.
Predatoren ... 20
6.1.2.
Detritivoren ... 21
6.1.3.
Mieren ... 22
Is het microklimaat een intermediaire factor voor de invloed van het landschap op de
activiteit-densiteit van arthropoda? ... 22
7. Algemeen besluit ... 23
8. Referentielijst ... 24
9. Bijlagen ... 29
Indeling van de ecosysteemdiensten volgens CICES ... 29
Scree plot van de principale componenten ... 30
1
1. LIJST MET TABELLEN EN FIGUREN
Lijst met tabellen
Tabel 1: Overzicht van welk habitattype in welke landschapscategorie werd ondergebracht en de verklaring van het habitattype (Loos, 2019). ... 12 Tabel 2: p-waarden van de Mann-Whitney U testen. De landschapsvariabelen werden opgesplitst in absence en presence groepen bij miljoenpoten en duizendpoten. ... 16 Tabel 3: Correlatiecoëfficiënten van de getransformeerde activiteit-densiteit van de arthropodagroepen en landschapsvariabelen op 10, 50 en 500 meter. Significante correlatiecoëfficiënten worden met een * weergegeven. ... 17 Tabel 4: waarden van de F-test weergegeven voor de vier modellen van alle arthropodagroepen. De p-waarden werden afgerond naar 2 cijfers na de komma en significante p-p-waarden zijn met een * gemarkeerd. ... 17 Tabel 5: Regressiemodellen van de zeven arthropodagroepen met de landschapsvariabelen binnen een straal van 50 m. Voor ieder model worden de R2-waarde, de schatter voor het intercept (B0) en de predictoren
in volgorde van toevoeging met hun ongestandaardiseerde coëfficiënt (B) weergegeven. Significante coëfficiënten worden met een * gemarkeerd (p < 0,05). ... 17 Tabel 6: Correlatiecoëfficiënten van activiteit-densiteit van de arthropodagroepen met het gemiddelde en de variantie van de temperatuur. Correlatiecoëfficiënten die significant verschillend zijn van 0 werden gemarkeerd met een *. ... 19 Tabel 7: Overzicht van de indeling van biotische ecosysteemdiensten bij CICES V5.1 voor de drie hoogste niveaus. ... 29 Tabel 8: Normaliteitstesten (Kolmogorov-Smirnov en Shapiro-wilk toets) voor het nagaan van de voorwaarden bij de opgebouwde regressiemodellen. De significante p-waarden werden met een * gemarkeerd. ... 31
Lijst met figuren
Figuur 1: Indeling van ecosysteemdiensten volgens het MA (2003). Er worden vier categorieën onderscheiden: voorziend (Provisioning Services), regulerend (Regulating Services), cultureel (Cultural Services) en ondersteunend (Supporting Services) (MA, 2003). ... 4 Figuur 2: twee hypothesen over de relatie tussen biodiversiteit en ecosysteemfuncties. Het aantal soorten in een ecosysteem (biodiversiteit) in functie van het aantal soorten dat ecosysteemfuncties levert. (a) Er zijn veel soorten met dezelfde ecosysteemfuncties. Zolang het verlies aan soorten beperkt blijft, is er geen verlies aan ecosysteemfuncties. (b) Zelfs zeldzame soorten hebben specifieke ecosysteemfuncties. Een verlies van soorten zorgt voor een verlies aan ecosysteemfuncties (Schröter et al., 2019). ... 5 Figuur 3: Relatieve impact van processen die een bedreiging vormen voor diersoorten in de Verenigde Staten. Opmerking: percentages gaan niet tot 100% omdat verschillende soorten door meerdere processen bedreigd worden (Lindenmayer & Fischer, 2006). ... 8 Figuur 4: Direct en indirect effect van landschapssimplificatie op de uiteindelijke gewasopbrengst door veranderingen in soortenrijkdom van natuurlijke vijanden (richness), de populatiegrootte (abundance), en natuurlijke plaagbestrijding (pest control) (alleen insecticidevrije percelen werden beschouwd, N = 184 percelen bij 14 gewassen) (Dainese et al., 2019). ... 9
2 Figuur 5: foto van het meettuintje Rooberg, genomen op 11 juli 2018 door tuinierster Martine Braeckman. In het tuintje staan radijs, spruitkool, andijvie, Chinese kool, rucola, bladpeterselie, plantui, knolvenkel, kropsla, zomerprei en aardbei volgens een vast plantschema. De bodemval (A) en de bodemtemperatuursensor (B) worden aangeduid op de foto. ... 11 Figuur 6: De landschapssamenstelling werd bepaald binnen concentrische cirkels van 10, 50 en 500 meter rondom de tuintjes. Hier wordt tuintje 33 (Proefhoevestraat, links) weergegeven als voorbeeld van een tuintje met hoofdzakelijk landbouw en tuintje 22 (Natuurpunt, rechts) als voorbeeld van een tuintje met hoofdzakelijk laag groen. ... 11 Figuur 7: Gemiddelde en standaarddeviatie van de activiteit-densiteit per arthropodagroep weergegeven voor de drie vangstperiodes (juni, juli en augustus). ... 13 Figuur 8: Loading plot van de arthropodagroepen met componenten 1 en 2 die samen 55,4% van de totale variantie, verklaren, waarbij component 1 38,2% verklaart en component 2 17,2%. Elke arthropodagroep wordt geplot met als coördinaten de loadings op de eerste twee principale componenten... 15 Figuur 9: Scatterplot van de scores voor component 1 en component 2 waarbij de datapunten van de tuintjes ingedeeld worden volgens landschapssamenstelling op a) 10, b) 50 en c) 500 meter. De tuintjes werden opgedeeld in HG (hoog groen), LB (landbouw), LG (laag groen) of NG (niet groen) volgens het grootste aandeel. ... 15 Figuur 10: Aandeel van de vier landschapsgroepen (landbouw, hoog groen, laag groen en niet groen) op 50 meter bij absence (blauw) en presence (oranje) van a) miljoenpoten en b) duizendpoten. De vier landschapsgroepen werden elk opgedeeld in twee subgroepen die werden bepaald door absence en presence van a) miljoenpoten en b) duizendpoten. Vervolgens werd de verdeling van deze subgroepen gevisualiseerd aan de hand van boxplots bij de vier landschapsgroepen. ... 16 Figuur 11: Boxplots van alle temperatuursmetingen uit 2019 weergegeven per tuintje. Voor de indeling volgens landschap werd de landschapscategorie bepaald met het grootste aandeel op 10, 50 en 500 meter voor alle tuintjes. De finale landschapsgroep voor een tuintje is deze die 2 of 3 keer het grootste aandeel had. Wanneer een tuintje 3 verschillende landschapsgroepen heeft bij 10, 50 en 500 meter, worden deze allemaal weergegeven. LG (lichtgroen): laag groen tuintje, HG (donkergroen): hoog groen tuintje, LB (geel): landbouw tuintje, NG (grijs): niet groen tuintje, rood: ander. ... 18 Figuur 12: Scree plot van de principale componenten die de variabiliteit verklaren van de activiteit-densiteit bij de arthropodagroepen. Bij de tweede component is er een knik te zien, waardoor ervoor wordt gekozen om twee componenten te behouden. ... 30 Figuur 13: scatterplots van de ongestandaardiseerde residuen in functie van de ongestandaardiseerde voorspelde waarden weergegeven voor de regressiemodellen van a) spinnen b) miljoenpoten c) loopkevers d) kortschildkevers e) mieren f) duizendpoten. ... 31
3
2. INLEIDING
Veranderingen in landgebruik hebben sinds de twintigste eeuw gezorgd voor achteruitgang van de habitatkwaliteit. Dit heeft kwalijke gevolgen voor de biodiversiteit en daaraan verbonden ecosysteemdiensten (von Haaren et al., 2019). In deze thesis worden functionele tuintjes gebruikt als meetinstrument voor de ecosysteemdiensten langs een landschappelijke complexiteitsgradiënt op het platteland. De ecosysteemdienst die hier nader onderzocht wordt, is natuurlijke plaagbestrijding. Dit gebeurt aan de hand van zijn leveranciers, de natuurlijke vijanden. Er wordt onderzocht wat het effect is van de landschapssamenstelling op de activiteit-densiteit van in de grond kruipende arthropoda. Er wordt ook exploratief onderzocht of het microklimaat hierbij een intermediaire factor kan zijn. De arthropoda uit de functionele groep ‘natuurlijke vijanden’ die hier onderzocht worden zijn loopkevers, kortschildkevers, duizendpoten en spinnen. De vangstmethode laat toe dat ook arthropoda uit andere functionele groepen verzameld worden waarvan er enkele meegenomen worden bij de bespreking van de resultaten. Zo zullen ook detritivoren (miljoenpoten, pissebedden) en mieren worden besproken.
In de literatuurstudie worden eerst ecosysteemdiensten besproken, om vervolgens toe te spitsen op natuurlijke plaagbestrijding en deze te bespreken aan de hand van zijn leveranciers, de natuurlijke vijanden. In de laatste sectie van de literatuurstudie wordt besproken welke invloed het landschap heeft bij de levering van ecosysteemdiensten. Hoe het onderzoek werd uitgevoerd, wordt toegelicht in het onderdeel ‘materiaal en methoden’. Het onderdeel ‘resultaten’ geeft weer wat de bevindingen zijn van het onderzoek, deze bevindingen worden toegelicht en besproken in het onderdeel ‘discussie’. In het onderdeel ‘algemeen besluit’ worden de belangrijkste bevindingen weergegeven over de invloed van het landschap op de activiteit-densiteit van de arthropoda.
4
3. LITERATUURSTUDIE
Ecosysteemdiensten
3.1.1. Definitie en indeling van ecosysteemdiensten
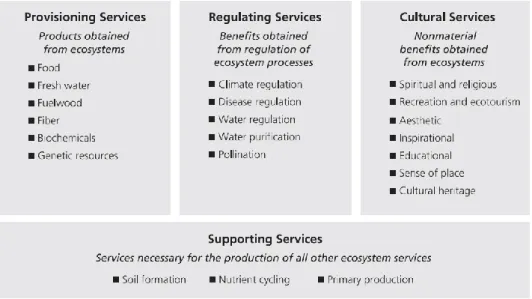
De meest gebruikte definitie voor ecosysteemdiensten is deze van het MA (Millenium Ecosystem Assessment): ecosysteemdiensten zijn de voordelen die mensen halen uit ecosystemen (MA, 2003). Het MA classificeert ecosysteemdiensten in 4 categorieën: voorziend, regulerend, cultureel en ondersteunend (figuur 1). Tot de voorzieningsecosysteemdiensten behoren voedselvoorziening en voorziening van natuurlijke bronnen die afkomstig zijn van ecosystemen. Regulerende ecosysteemdiensten zijn de voordelen verkregen door de regulatie van processen in het ecosysteem. Culturele ecosysteemdiensten zijn de niet-materiële voordelen die mensen verkrijgen van ecosystemen door spirituele verrijking, cognitieve ontwikkeling, reflectie, recreatie en esthetische ervaringen. Ondersteunende diensten zijn nodig voor de productie van alle andere ecosysteemdiensten. In tegenstelling tot de eerder vernoemde ecosysteemdiensten hebben ondersteunende ecosysteemdiensten een indirect voordeel voor de mens of is er een lange termijn effect (MA, 2003).
Figuur 1: Indeling van ecosysteemdiensten volgens het MA (2003). Er worden vier categorieën onderscheiden: voorziend (Provisioning Services), regulerend (Regulating Services), cultureel (Cultural Services) en ondersteunend (Supporting Services) (MA, 2003).
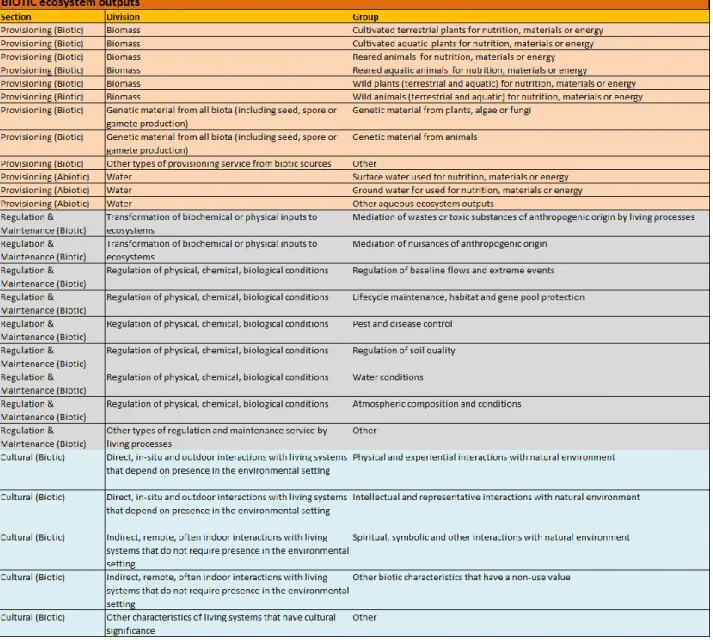
Een recentere indeling van ecosysteemdiensten gebeurde door CICES (Common International Classification of Ecosystem Services). Deze organisatie heeft als doel om enige standaardisering te bekomen in de manier waarop ecosysteemdiensten beschreven worden. De eerst bruikbare versie van CICES (V4.3) werd beschikbaar in 2013, de meest recente versie, CICES 5.1, is beschikbaar sinds 2018 (CICES). CICES maakt gebruik van een hiërarchische structuur, het hoogste niveau bestaat uit secties die geïntroduceerd werden door het MA. Regulerende en onderhoudende ecosysteemdiensten worden echter binnen dezelfde sectie beschouwd. Er zijn dus 3 secties: voorzienings-, regulerende en onderhoudende, en culturele ecosysteemdiensten (Haines-Young et al., 2012). CICES 5.1 beschrijft 5 niveaus: sectie, verdeling, groep, klasse, klassensoort. Een overzicht van de indeling van de biotische ecosysteemdiensten is te vinden in tabel 8 in bijlage 9.1 (Haines-Young et al., 2012).
5 In deze masterproef is voornamelijk de regulerende ecosysteemdienst die zorgt voor natuurlijke plaagbestrijding van belang. Natuurlijke plaagbestrijding is een ecosysteemdienst waarbij veranderingen in het ecosysteem zorgen voor minder plagen en ziektes bij vee en gewassen (MA, 2003). In volgende paragrafen wordt deze ecosysteemdienst en zijn leveranciers, natuurlijke vijanden, verder besproken. Er wordt eveneens bestudeerd welke invloed het landschap uitoefent op deze leveranciers, de nadruk ligt hierbij op het platteland. Landbouwers zijn afhankelijk van ecosysteemdiensten voor hun productie. Zo zorgt natuurlijke plaagbestrijding ervoor dat plagen minder schade aanrichten en de opbrengst aldus verhoogd wordt. Deze belangrijke ecosysteemdienst wordt echter in toenemende mate bedreigd door verlies aan biodiversiteit, modernisering van de landbouw en door menselijke invloed op natuurlijke ecosystemen (Zhang et al., 2007).
3.1.2. Ecosysteemdiensten en biodiversiteit
Kremen (2005) definieert ecosysteemdiensten als ‘het deel van de ecosysteemfuncties dat nuttig is voor de mens’. Deze ecosysteemfuncties zijn sterk gerelateerd aan de biodiversiteit van het ecosysteem; onderzoek over biodiversiteit en ecosysteemfuncties toont aan dat door een extreem verlies aan biodiversiteit de stabiliteit van het ecosysteem en het aantal ecosysteemfuncties gereduceerd wordt (Schröter et al., 2019). Er is echter nog steeds onduidelijkheid hoe biodiversiteit verschillende ecosysteemfuncties (en dus ecosysteemdiensten) ondersteunt (Schröter et al., 2019). Het is onbekend of er bijvoorbeeld enkele dominante soorten of veel complementaire soorten nodig zijn om ecosysteemdiensten te voorzien (Dainese et al., 2019). Verschillende onderliggende factoren kunnen de respons van ecosysteemdiensten op biodiversiteit veranderen. Bovendien is interactie tussen deze factoren mogelijk. Zo hebben enerzijds gemeenschappen met een grote soortenrijkdom meer kans om soorten te bevatten die verantwoordelijk zijn voor grootschalige effecten op het ecosysteem. Anderzijds kunnen dergelijke diverse ecosystemen een specifieke soortencombinatie bevatten waarbij de soorten complementair zijn (Dainese et al., 2019). Er zijn twee hypothesen over de relatie tussen het aantal soorten in een ecosysteem (de biodiversiteit) en het aantal soorten dat ecosysteemfuncties levert (Fig. 2). De eerste hypothese stelt dat er veel soorten zijn met dezelfde ecosysteemfuncties. Verlies van het aantal soorten veroorzaakt geen verlies van ecosysteemfuncties zolang er grote functionele groepen aanwezig zijn. Iedere functionele groep bestaat hierbij uit soorten die eenzelfde ecosysteemfunctie leveren. In de tweede hypothese gaat men ervan uit dat zelfs zeldzame soorten specifieke ecosysteemfuncties hebben. Hier resulteert een verlies van soorten onvoorwaardelijk in een verlies van ecosysteemfuncties (Schröter et al., 2019).
(a) (b)
Figuur 2: twee hypothesen over de relatie tussen biodiversiteit en ecosysteemfuncties. Het aantal soorten in een ecosysteem (biodiversiteit) in functie van het aantal soorten dat ecosysteemfuncties levert. (a) Er zijn veel soorten met dezelfde ecosysteemfuncties. Zolang het verlies aan soorten beperkt blijft, is er geen verlies aan ecosysteemfuncties. (b) Zelfs zeldzame soorten hebben specifieke ecosysteemfuncties. Een verlies van soorten zorgt voor een verlies aan ecosysteemfuncties (Schröter et al., 2019).
6
Arthropoda
als leveranciers van ecosysteemdiensten:
natuurlijke vijanden
In deze thesis wordt verder ingegaan op één ecosysteemdienst: natuurlijke plaagbestrijding. Deze ecosysteemdienst wordt verder besproken aan de hand van zijn leveranciers: natuurlijke vijanden. De relatie tussen natuurlijke vijanden en plaagbestrijding wordt hier niet nader toegelicht. Natuurlijke vijanden van een soort zijn organismen die deze soort doden, het reproductievermogen van deze soort verminderen, of het aantal organismen van deze soort reduceren (Flint & Dreistadt, 1998). Natuurlijke vijanden zorgen voor natuurlijke plaagbestrijding in een agro-ecosysteem. Er zijn twee functionele groepen die de ecosysteemdienst natuurlijke plaagbestrijding kunnen leveren: predatoren en parasieten (Dainese et al., 2019). Predatoren of roofdieren zijn organismen die één of meerdere soorten (de prooi) aanvallen, doden en als voedsel gebruiken (Flint & Dreistadt, 1998). Parasieten zijn organismen die leven ten koste van een ander organisme, de gastheer (Nederlandse encyclopedie). De natuurlijke vijanden die in de literatuur worden besproken zijn vertegenwoordigers van de taxonomische ordes Coleoptera (kevers), Diptera (tweevleugeligen), Aranea (spinnen), Hymenoptera (vliesvleugeligen), Hemiptera (halfvleugeligen), Thysanoptera (thrips) en Neuroptera (netvleugeligen) (Dainese et al., 2019). Hieronder worden de arthropoda besproken die belangrijke natuurlijke vijanden zijn en relevant zijn voor het praktisch werk van deze thesis.
3.2.1. Coleoptera
Alle kevers en hun larven hebben bijtende monddelen (Bellman, 2018). De orde van de kevers bevat dan ook veel families die voornamelijk uit predatoren bestaan. Lieveheersbeestjes en loopkevers worden het vaakst ingezet als natuurlijke vijand in land- en tuinbouwbedrijven (Debach & Rosen, 1991).
Cantharidae: weekschildkevers
De weekschildkevers hebben behoorlijk zachte dekschilden met een kleur die sterk varieert binnen de soort (Bellman, 2018). Adulten leven van pollen en nectar, meestal zijn ze ook predatoren van zachte insecten. De larven zijn hoofdzakelijk verantwoordelijk voor de natuurlijke plaagbestrijding. Ze jagen voornamelijk op insecten met een weke huid en bestrijden plagen aldus voornamelijk in het larvale stadium (Traugott, 2002).
Carabidae: loopkevers
Loopkevers kunnen carnivoor, herbivoor of omnivoor zijn. Het voedingsgedrag kan afhankelijk zijn van het seizoen en de beschikbaarheid van hun prooi. 73,5% Van de soorten uit de familie van de Carabidae en Cicineliae is uitsluitend carnivoor; 8,1% is fytofaag en 19,5% is omnivoor (Larochelle, 1990). Bijna alle leden van de loopkever familie zijn rovers van andere insecten en wormen (Bellman, 2018). Ze zijn predatoren van luizen, wormen, cicaden, rupsen, slakken, springstaarten en verschillende insectenlarven (Lövei & Sunderland, 1996). Deze kevers overwinteren in halfnatuurlijke landschapselementen zoals houtkanten en grasranden. In het begin van het groeiseizoen verplaatsen ze zich naar het landbouwperceel waar ze de eerste generaties plaaginsecten bestrijden (Pfiffner & Luka, 2000).
Coccinellidae: lieveheersbeestjes
Bijna alle lieveheersbeestjes hebben een breed-ovaal, nagenoeg cirkelvormig lichaam. Deze familie bestaat grotendeels uit carnivoren waarbij het voornaamste voedsel van larven en adulten uit bladluizen bestaat. Een kleine minderheid voedt zich met planten (Bellman, 2018; Lundgren, 2009). Enkele soorten uit deze familie zijn predatoren van schildluizen, bladhaantjes, tripsen, vlindereitjes of mijten. Voor lieveheersbeestjes is het belangrijk dat er ook niet-prooi voedsel aanwezig is onder de vorm van nectar, honingdauw, stuifmeel, fruit, vegetatie en schimmel. Deze alternatieve voeding wordt gebruikt om de overlevingskansen te verhogen wanneer slechts kleine prooipopulaties beschikbaar zijn, om de sterfte tijdens de diapauze te verminderen, de brandstofmigratie te verminderen en de reproductiecapaciteit te verbeteren (Lundgren, 2009).
7
Staphylinidae: kortschidlkevers
Kortschildkevers hebben een slank lichaam en korte dekschilden die alleen de twee eerste abdomen segmenten bedekken (Bellman, 2018). Deze kevers hebben stekende-zuigende monddelen, ze maken eerst een opening in hun prooi waarlangs speeksel toegediend wordt om de opgeloste inhoud vervolgens op te zuigen (Betz et al., 2018). Veel kortschildkevers zijn predatoren en voeden zich met andere arthropoda, eitjes en larven van vliegen, rupsen, slakken, bladluizen, mijten of andere kleine organismen (Van Driesche & Bellows, 1996; Betz et al., 2018). Kortschildkevers kunnen ook (obligaat of facultatief) fungivoor, herbivoor of saprofyt zijn (Betz et al., 2018).
3.2.2. Araneae
De orde van de spinnen (Araneae) bestaat uitsluitend uit predatoren die leven van onder andere trips, rupsen, bladluizen, insecten, sprinkhanen en vliegen (Van Driesche & Bellows, 1996; Cushing, 2008). De prooi kan gevangen worden op verschillende manieren. Er zijn soorten die hun prooi vangen in een web, andere soorten jagen actief op hun prooi. Na de vangst wordt gif geïnjecteerd en wordt de prooi ingewikkeld met spinsel (Cushing, 2008). Spinnen zijn weinig kieskeurig wat hun prooi betreft maar vertonen een sterke habitat specificiteit. Deze plaagbestrijders zijn dus weinig geschikt voor introductie bij het onder controle houden van plagen in nieuwe gebieden. Wanneer spinnen dus gewenst zijn als natuurlijke vijanden, gebeurt dit best door het bevoordelen van gebiedseigen spinnen via cultuurtechnische maatregelen (Van Driesche & Bellows, 1996).
3.2.3. Hymenoptera
Alle vliesvleugeligen (orde Hymenoptera) hebben bijtende monddelen, maar adulten leven van vloeibaar voedsel. Er zijn twee subordes: blad-, houten halmwespen en verwanten (Symphyta) en sluipwespen, wespen, mieren en bijen (Apocrita) (Bellman, 2018). Voor natuurlijke plaagbestrijding zijn voornamelijk de sluipwespen (Parasitica) van belang. Sluipwespen hebben een legboor die ze gebruiken om gif in hun gastheer te injecteren. Dit gif heeft een verdovende werking zodat ze eitjes in, op of naast de gastheer kunnen leggen. Wanneer de eitjes uitkomen, ontwikkelen de larven zich ten koste van deze gastheer (Bellman, 2018).
8
Invloed van het landschap op natuurlijke vijanden
3.3.1. Belang van het landschap voor natuurlijke vijanden
De natuurlijke vijanden die eerder besproken werden zijn afhankelijk van habitats voor het leveren van hun ecosysteemdienst (natuurlijke plaagbestrijding). Deze habitats bevatten voorzieningen zoals voedsel, schuilplaats tegen predatoren en overwinteringsplaatsen (Kremen et al., 2007; Mitchell et al. 2014). De habitat van natuurlijke vijanden is echter vaak in tijd of ruimte gescheiden van de locatie waar de mens wil dat ze hun ecosysteemdienst leveren (Kremen et al., 2007). Het is dus noodzakelijk dat er in de omgeving landschapselementen aanwezig zijn die voor de natuurlijke vijanden als habitat kunnen dienen. De habitatkwaliteit is bepalend voor de aan- of afwezigheid van een soort (Duelli, 1997). Veranderingen in landgebruik hebben sinds de twintigste eeuw gezorgd voor achteruitgang van de habitatkwaliteit, dit resulteerde in een aanzienlijk verlies van ecosysteemdiensten (von Haaren et al., 2019). Natuurlijke habitat verdween of versnipperde, om plaats te maken voor gebouwen en landbouwakkers. Intensivering van de landbouw zorgde voor meer insecticidengebruik, wat het potentiële effect van natuurlijke plaagbestrijding reduceert. Plaagbestrijding is positief gecorreleerd met gewasopbrengst wanneer geen gebruik gemaakt wordt van insecticiden, in besproeide zones heeft de plaagbestrijding geen invloed op de gewasopbrengst (Dainese et al., 2019). Bij landen waar intensieve landbouw heeft geleid tot verontrustende achteruitgang van ecosystemen (en hun diensten), zijn er steeds meer inspanningen om de landbouwlandschappen te revitaliseren (Duelli, 1997). Er is dan ook veel onderzoek naar de rol van het landschap bij de levering van ecosysteemdiensten. Hier wordt de rol van het landschap besproken aan de hand van de voornaamste bedreigingen voor diersoorten in de Verenigde Staten: habitatverlies en -fragmentatie (Lindenmayer & Fischer, 2006) (Fig. 3).
3.3.2. Habitatfragmentatie
Habitatfragmentatie is het splitsen van een habitat in twee of meer kleinere gebieden. Deze verschillende kleinere habitatgronden zijn mogelijks te klein om levensvatbare populaties van een soort in stand te houden. Bovendien kan habitatfragmentatie ervoor zorgen dat de afstand tussen habitats groter wordt waardoor ze meer geïsoleerd zijn van elkaar (Lindenmayer & Fischer, 2006). De veranderingen in landgebruik die habitatfragmentatie veroorzaken, hebben nadelige gevolgen voor de landschapsconnectiviteit (Mitchell et al., 2013). Dit is de mate waarin een landschap de beweging van organismen en materie vergemakkelijkt (Crooks & Sanjayan, 2006; Mitchell et al., 2013). Het is afhankelijk van de structuur en compositie van het landschap (structurele connectiviteit) enerzijds, en van de respons van de organismen en materie op die structuur
Figuur 3: Relatieve impact van processen die een bedreiging vormen voor diersoorten in de Verenigde Staten. Opmerking: percentages gaan niet tot 100% omdat verschillende soorten door meerdere processen bedreigd worden (Lindenmayer & Fischer, 2006).
9 (functionele connectiviteit) anderzijds (Mitchell et al., 2013). Connectiviteit is van cruciaal belang voor de biodiversiteit, het is de bepalende factor voor uitwisseling van genen in een populatie. Bij connectiviteitsverlies kunnen soorten minder habitatpercelen bereiken en gebruiken, dit zal resulteren in een afname van het aantal verspreidingsgebieden en van de populatiegroottes (Mitchell et al., 2013; Herrera et al., 2017).
3.3.3. Habitatverlies en vermindering van de habitatkwaliteit
Vermindering van de habitatkwaliteit is de langzame achteruitgang van habitatgeschiktheid of de uitputting van habitats. Dit proces kan uiteindelijk leiden tot habitatverlies als het blijft evolueren en niet wordt stopgezet (Lindenmayer & Fischer, 2006). Schaalvergroting bij landbouwbedrijven resulteerde in vereenvoudiging van het landschap of landschapssimplificatie op het platteland. Bijgevolg is er op het platteland veel habitatverlies en vermindering van de habitatkwaliteit (Bianchi et al., 2006). Landschapssimplificatie houdt verband met de structuur van het landschap. Op het platteland omvat dit de configuratie en compositie van zowel gewas als niet-gewas habitat (Mitchell et al., 2014). Het heeft een effect op de beweging en diversiteit van natuurlijke vijanden en beïnvloedt aldus de natuurlijke plaagbestrijding (Bianchi et al., 2006; Chaplin-Kramer et al., 2011). De meeste studies die de relatie tussen het landschap en de natuurlijke plaagbestrijding bestuderen, hanteren landschapscomplexiteit als parameter voor de structuur van het landschap (Mitchell et al., 2014). Dit kan gemeten worden aan de hand van het aandeel niet-gewas habitat (Batáry et al., 2011), de diversiteit van aanwezige habitats (Fabian et al., 2013; Gardiner et al., 2009) of de aanwezigheid van lineaire elementen zoals bomenrijen (Holzschuh et al. 2010).
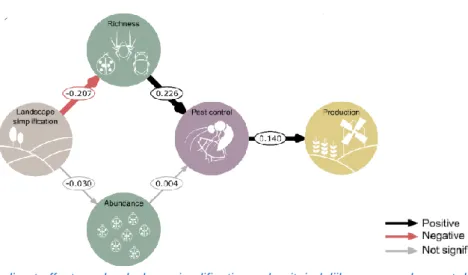
Op figuur 4 wordt aangetoond dat landschapssimplificatie negatieve gevolgen heeft voor natuurlijke plaagbestrijding door de soortenrijkdom (richness) van de natuurlijke vijanden te benadelen, en niet zozeer via de populatiegrootte (abundance) (Dainese et al., 2019). In sectie 1.2 werd het belang van biodiversiteit voor het leveren van ecosysteemdiensten reeds vermeld, natuurlijke plaagbestrijding is zo een ecosysteemdienst die aangedreven wordt door biodiversiteit. Hoe meer verschillende soorten predatoren er dus aanwezig zijn in een ecosysteem, hoe meer natuurlijke plaagbestrijding er zal plaatsvinden (Dainese et al., 2019). Predatorgroepen met een grotere biodiversiteit zullen namelijk plaaginsecten op meer verschillende manieren bestrijden in tijd en ruimte, waardoor de natuurlijke plaagbestrijding effectiever zal verlopen (Mitchell et al., 2014). Bovendien is een ‘sampling effect’ mogelijk; hoe groter de soortenrijkdom van een populatie, hoe meer kans dat hierin de meest effectieve natuurlijke vijand van een herbivoor aanwezig is (Tscharntke et al., 2005).
Figuur 4: Direct en indirect effect van landschapssimplificatie op de uiteindelijke gewasopbrengst door veranderingen in soortenrijkdom van natuurlijke vijanden (richness), de populatiegrootte (abundance), en natuurlijke plaagbestrijding (pest control) (alleen insecticidevrije percelen werden beschouwd, N = 184 percelen bij 14 gewassen) (Dainese et al., 2019).
10
4. MATERIAAL EN METHODEN
Dit onderzoek maakt deel uit van het ‘BEL-Landschap’ project (Biodiversiteit, Ecosysteemdiensten en Landbouw). Het project heeft als doel het belang van half-natuurlijke elementen in het landschap te kwantificeren d.m.v. moestuintjes die het effect van de (biodiversiteit in de) omgeving op de gewassen in de tuintjes opmeten. Het project liep over twee groeiseizoenen, 2018 en 2019. In dit deel van het project worden uitsluitend de arthropoda data van 2019 gebruikt. In 2018 werden er 40 identieke moestuintjes verspreid over het onderzoeksgebied, in 2019 waren er nog 38 tuintjes. Het gekozen onderzoeksgebied is representatief voor peri-urbaan Vlaanderen en bevindt zich in Gontrode, Landskouter, Gijzenzele en Lemberge. 90 Vrijwilligers hielpen bij het onderhoud van de tuintjes en de dataverzameling, ze werden op de hoogte gehouden via een wekelijkse nieuwsbrief (https://www.bel-landschap.be/). Om dit onderzoek uit te kunnen voeren, werden gegevens gebruikt over de arthropoda, het microklimaat en de landschapssamenstelling. In volgende secties wordt verduidelijkt hoe deze gegevens verzameld en verwerkt werden.
Dataverzameling van de arthropoda en het microklimaat via
functionele meettuintjes
4.1.1. Meetinstrument: functionele tuintjes
De meettuintjes hebben een grootte van 1 m2 en werden omheind door kippengaas om de gewassen te
beschermen tegen wildvraat. De tuinbakken werden gevuld met groeisubstraat. In 2018 was dit standaard moestuingrond, namelijk TerraViva® Moestuingrond. In 2019 was het substraat een mengsel van compost en volle grond die afgegraven werd van een proefperceel op het ILVO in een optimale verhouding van 3:7 (https://www.bel-landschap.be/). Er werden ongeveer 10 gewassen geteeld volgens een vaststaand schema: kropsla, knolvenkel, andijvie, Chinese kool, radijs, spruitkool, plantui, rucola, peterselie, aardbei, zomerprei (Fig. 5). Opdat de tuintjes een geschikt meetinstrument zouden vormen, werden ze door de vrijwilligers zoveel mogelijk op uniforme wijze behandeld (watergift, oogst, onkruidbeheersing). Dit werd mogelijk gemaakt door een nieuwsbrief die wekelijks werd uitgestuurd naar de vrijwilligers en dankzij een draaiboek met gewasfiches.
4.1.2. Dataverzameling van de arthropoda
De grond-arthropoda werden verzameld via bodemvallen. Deze methode maakt het mogelijk om de fractie natuurlijke plaagbestrijders te vangen die in de grond kruipen. In ieder tuintje werd een bodemval ingegraven en gevuld met een mengsel van antivries en detergent. De druppel detergent zorgt ervoor dat de arthropoda niet op het vloeistofoppervlak blijven drijven door de oppervlaktespanning (Luff & Topping,1995). De val werd voorzien van een afdak om te voorkomen dat deze gevuld werd met gietwater of regenwater (Fig. 5A). Na twee weken werden deze vallen opgehaald. Deze procedure werd in 2018 en 2019 drie keer uitgevoerd; in juni, juli en augustus. Alle dieren in de vallen werden ondergebracht in één van de volgende groepen: spinnen (Araneae), pissebedden (Isopoda), miljoenpoten (Diplopoda), duizendpoten (Chilopoda), loopkevers (Carabidae), kortschildkevers (Staphylinidae), mieren (Formicidae), andere (bv. lieveheersbeestjes, slakken, hooiwagens…).
4.1.3. Dataverzameling van het microklimaat
In het midden van ieder tuintje werd een bodemtemperatuursensor HOBO Pendant T® (8k) geplaatst (Fig. 5B). Deze sensor heeft ieder uur de temperatuur tijdens het groeiseizoen (van 29/05/2019 t.e.m. 28/09/2019). De sensor heeft een meetbereik van -20° tot 70°C en een accuraatheid van ± 0.53°C bij temperaturen tussen 0° en 50°C (Loos, 2019).
11
Landschapssamenstelling 10, 50 en 500 meter rondom de
tuintjes
Bij het bepalen van de locatie van de tuintjes werd een goede verdeling over een gradiënt van het percentage half natuurlijke elementen (hoog groen en laag groen) in de omgeving beoogd (https://www.bel-landschap.be/NL/Informatie/Landschapsobservatorium/). Hoewel ieder tuintje toegankelijk was voor mensen of zich in de nabije omgeving van menselijke woningen bevond, is landschapssamenstelling rondom de tuintjes is zeer gevarieerd. In 2018 werd rondom ieder tuintje de samenstelling van het landschap bepaald binnen concentrische cirkels rondom het tuintje met een straal van 10, 50 en 500 meter (Loos, 2019). Bij dit onderzoek wordt deze dataset gebruikt. Er zijn vier landschapscategorieën: landbouw, hoog groen, laag groen en niet groen (Fig. 6) waarin verschillende habitattypes kunnen worden ondergebracht (Tabel 1). De landschapssamenstelling binnen een straal van 10 meter wordt gebruikt om de invloed van de directe omgeving te onderzoeken. De categorieën werden bepaald via een visuele inschatting tijdens een veldbezoek in 2018. De afstand van 50 meter geeft een idee over de invloed van de ruimere omgeving. De afstand van 500 meter werden gekozen om een inschatting te geven van de landschapssamenstelling rondom het tuintje op grote schaal, zonder dat de concentrische cirkels van de meettuintjes elkaar in grote mate zouden overlappen (Loos, 2019). De landschapssamenstelling binnen deze concentrische cirkels werden bepaald via het programma ArcGis. Hiertoe werden de locaties van de meettuintjes eerst als punten ingetekend a.d.h.v. een recente luchtfoto. Vervolgens werden cirkels met een straal van 10, 50 en 500m rondom de meettuintjes bepaald en werden aparte shapefiles per straal gecreëerd. Daarna werd een habitatkaart gemaakt voor elk van de vooraf bepaalde habitattypes, deze habitatkaart werd omgezet naar rasterformaat indien nodig. Tenslotte kon de oppervlakte van de habitattypes bepaald worden binnen de concentrische cirkels (Loos, 2019).
Figuur 6: De landschapssamenstelling werd bepaald binnen concentrische cirkels van 10, 50 en 500 meter rondom de tuintjes. Hier wordt tuintje 33 (Proefhoevestraat, links) weergegeven als voorbeeld van een tuintje met hoofdzakelijk landbouw en tuintje 22 (Natuurpunt, rechts) als voorbeeld van een tuintje met hoofdzakelijk laag groen.
Figuur 5: foto van het meettuintje Rooberg, genomen op 11 juli 2018 door tuinierster Martine Braeckman. In het tuintje staan radijs, spruitkool, andijvie, Chinese kool, rucola, bladpeterselie, plantui, knolvenkel, kropsla, zomerprei en aardbei volgens een vast plantschema. De bodemval (A) en de bodemtemperatuursensor (B) worden aangeduid op de foto.
12
Tabel 1: Overzicht van welk habitattype in welke landschapscategorie werd ondergebracht en de verklaring van het habitattype (Loos, 2019).
Landschaps-categorie
Type habitat Verklaring
Landbouw Tuinbouw Landbouwpercelen bedekt met tuinbouwgewassen Akkerbouw Landbouwpercelen bedekt met akkerbouwgewassen
Hoog groen (vegetatie hoger dan 3 m)
Tuin hoog groen Bomen en struiken hoger dan 3 m die zich in een tuin bevinden. Urbaan hoog groen Bomen en struiken hoger dan 3 m die zich in een urbane
omgeving bevinden (uitgezonderd in tuin). Agrarisch hoog
groen
Bomen en struiken hoger dan 3 m die zich in een agrarische omgeving bevinden. Bv. alleenstaande bomen, bomenrijen, houtkanten hoger dan 3 m en bomengroepen.
Bos Hoog groen met een aaneengesloten kroonoppervlakte van groter dan 5000 m2.
Laag groen (vegetatie lager dan 3 m)
Moestuin Oppervlakte in de tuin, gebruikt om (eenjarige en vaste) gewassen op te telen.
Gazon Frequent gemaaide gras-kruidige vegetatie in de tuin. Houtig laag groen Lage vegetatiestroken in de tuin die niet ingezaaid werden. Bloembedden Oppervlakte in de tuin die ingezaaid of beplant zijn met
sierplanten omwille van de bloemen.
Bloemenstrook Lage vegetatiestroken die ingezaaid werden met een bloemenmengsel.
Gras-kruidenstrook Lage vegetatiestroken die niet ingezaaid werden.
Grasland Oppervlaktes bedekt met gras-kruidige lage vegetatie, in stand gehouden door begrazing, maaien of een combinatie.
Struweel, struiken en hagen
Houtige vegetatie lager dan 3 m (maar die zich niet in een tuin bevindt).
Niet groen Gebouwen Bebouwde oppervlakte
Verharding Grond bedekt met verharding (asfalt, geplaveid, kiezelstenen…) Water Waterlichamen zoals poelen, rivieren, meren enz.
Onbedekte bodem Onverharde ondergrond zonder vegetatiebedekking.
Dataverwerking van de resultaten
Voor de verwerking van de data wordt gebruik gemaakt van het programma ‘SPSS statistics 25’.
4.3.1. Transformatie van de arthropoda activiteit-densiteit
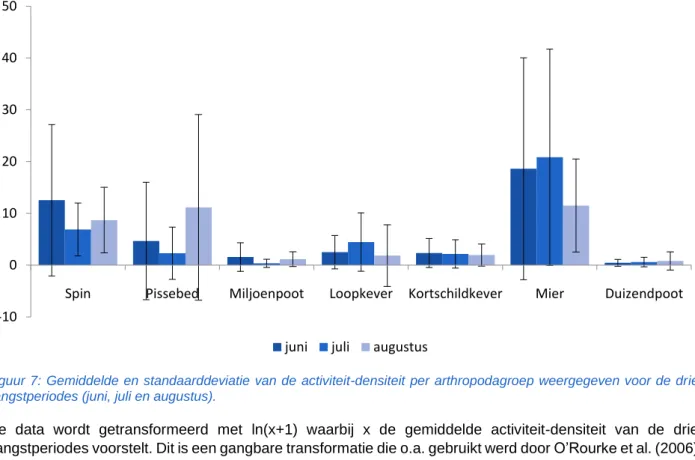
De hoeveelheid arthropoda die in een bodemval gevangen wordt is afhankelijk van zowel de activiteit als de densiteit van de lokale arthropodapopulaties. Data afkomstig van bodemvallen verwijst bijgevolg naar een waarde die bekend staat als ‘activiteit-densiteit’ (Thomas et al., 1998; Djoudi et al., 2018). De verdere bespreking van de data zal gebeuren met het gemiddelde van de drie vangstperiodes per tuintje. Er zijn geen duidelijke verschillen zichtbaar tussen de drie vangstperiodes (Figuur 7). De invloed van de vangstperiode op de activiteit-densiteit van een arthropodagroep wordt hier niet onderzocht.
13 De data wordt getransformeerd met ln(x+1) waarbij x de gemiddelde activiteit-densiteit van de drie vangstperiodes voorstelt. Dit is een gangbare transformatie die o.a. gebruikt werd door O’Rourke et al. (2006) en Flórián et al. (2019). Transformatie wordt vaak aangeraden om niet-normaal verdeelde fouttermen te vermijden waardoor de data in een lineair model toegepast kan worden. Bovendien kunnen transformaties het effect van outliers wegwerken en de spreiding over verschillende niveaus van een factor egaliseren, waardoor de lineariteit van de responsvariabele en de homoscedasticiteit verbeteren (St-Pierre et al., 2018). Er zijn echter nadelen verbonden aan deze methode en O’Hara & Kotze (2010) raden af om data logaritmisch te transformeren. Volgens St-Pierre et al. (2018) is deze methode empirisch: hij kan niet gebruikt worden in biologische vergelijkingen met gekende parameters. Ook het probleem van niet-lineaire vergelijkingen, zoals hyperbolische en asymptotische vergelijkingen, kan met een transformatie niet worden opgelost. Bovendien bekomt men door de transformatie moeilijk te interpreteren parameters omdat ze een schaal bezitten die verschilt van de oorspronkelijke gegevens (St-Pierre et al., 2018). Ives (2015) toont echter aan dat de type I fout het best vermeden kan worden door de data te transformeren. Ook St-Pierre et al. (2018) raden het gebruik van transformatie aan wanneer het vermijden van de type I-fout belangrijker is dan het toepassen van de oorspronkelijke schaal van een parameter. Bij dit onderzoek wordt ervoor gekozen om het risico op een type I-fout zo laag mogelijk te houden en de gemiddelde activiteit-densiteit te transformeren.
4.3.2. PCA-analyse
In deze thesis beschikken we over een multivariate dataset met verschillende arthropodagroepen. Er wordt dus eerst een exploratieve data-analyse uitgevoerd m.b.v. PCA op basis van de correlatiematrix. Eerst wordt de ‘Bartlett’s test of sphericity’ uitgevoerd om na te gaan of de correlatiematrix geen eenheidsmatrix is en of het bijgevolg zinvol is om de PCA uit te voeren (Constantin, 2014). Deze data-analyse wordt uitgevoerd bij de getransformeerde data om de invloed van outliers te reduceren (Venables & Ripley, 2002). De coördinaten van de tuintjes in het nieuwe PCA assenstelsel worden bepaald voor de eerste twee componenten en opgeslagen. Deze twee nieuwe variabelen zijn de scores van de tuintjes en kunnen voorgesteld worden op een scatterplot. De tuintjes worden uitgezet op de PCA volgens het type landschap met het grootste aandeel op 10, 50 en 500 meter.
Figuur 7: Gemiddelde en standaarddeviatie van de activiteit-densiteit per arthropodagroep weergegeven voor de drie vangstperiodes (juni, juli en augustus).
-10 0 10 20 30 40 50
Spin Pissebed Miljoenpoot Loopkever Kortschildkever Mier Duizendpoot
14
4.3.3. Opdeling in absence en presence data bij duizendpoten en miljoenpoten
Bij miljoenpoten en duizendpoten kwamen veel nulwaarden voor, in respectievelijk 27% en 24% van tuintjes werden deze arthropoda niet waargenomen. Dit zijn typische waarnemingen bij data afkomstig van bodemvallen (St-Pierre et al., 2018). De data van miljoenpoten en duizendpoten werd opgedeeld in een ‘absence’ en een ‘presence’ groep zodat onderzocht kan worden of ‘presence’ bepaald wordt door landschapssamenstelling. Er wordt alleen gewerkt met de landschapssamenstelling binnen een straal van 50 meter omdat we vermoeden dat deze nabije omgeving een significante invloed zal hebben op de gemeenschap. Volgens Thomas & Marshall (1999) is deze afstand geschikter voor het analyseren van data afkomstig van bodemvallen dan de zeer dichte omgeving van het meetpunt (10m). Bij de exploratieve data-analyse wordt visueel ingeschat d.m.v. boxplots of de landschapssamenstelling tussen de absence en presence groepen sterk verschilt. Vervolgens wordt een statistische test uitgevoerd om aan te tonen of er een significant verschil is in landschapssamenstelling tussen deze twee groepen. De voorwaarden voor One-way ANOVA zijn niet voldaan, er wordt gekozen voor de Mann-Whitney U test als niet-parametrisch alternatief.4.3.4. Meervoudige lineaire regressie
De data van miljoenpoten en duizendpoten wordt in deze sectie integraal verwerkt en dus niet meer opgedeeld in absence en presence groepen. Eerst worden er hypothesetesten uitgevoerd om na te gaan of er regressie is tussen x (het aandeel van de landschapsvariabele) en y (getransformeerde gemiddelde activiteit-densiteit). Vervolgens wordt er aan de hand van geautomatiseerde modelbouw onderzocht welke landschapsvariabelen een belangrijke invloed hebben op de activiteit-densiteit van de arthropoda. Om multicollineariteit te vermijden, worden de variabelen van slechts één van drie schalen gebruikt. Omdat we vermoeden dat de landschapssamenstelling binnen een straal van 50 meter het meest representatief zal zijn voor de omgeving van het tuintje, kiezen we ervoor om de landschapsvariabelen van deze schaal te gebruiken bij de opbouw van het regressiemodel. Het model wordt stapsgewijs opgebouwd uit de vier predictor-variabelen (hoog groen, laag groen, niet groen en landbouw) door middel van ‘forward selection’. Door deze methode wordt in de eerste stap de meest significante variabele toegevoegd. Bij elke volgende stap wordt de meest significante variabele toegevoegd die nog niet in het vorige model voorkwam, totdat alle variabelen zijn toegevoegd (Flom & Cassell, 2007). Het beste model wordt gekozen d.m.v. de F-test. Bij de regressiemodellen die bestaan uit meer dan één predictor, wordt onderzocht of de interactieterm significant is door deze toe te voegen aan het ‘general linear model’.
4.3.5. Exploratieve data-analyse bij de data van het microklimaat
De data van het microklimaat wordt eerst op boxplots voorgesteld, om vervolgens ingekleurd te worden volgens landschapscategorie. Voor de indeling volgens landschap werd de landschapscategorie bepaald met het grootste aandeel op 10, 50 en 500 meter voor alle tuintjes. De finale landschapsgroep voor een tuintje is deze die 2 of 3 keer het grootste aandeel had. Wanneer een tuintje 3 verschillende landschapsgroepen heeft bij 10, 50 en 500 meter, worden deze allemaal weergegeven. Voor het verband tussen het microklimaat en de arthropodagroepen werd voor ieder tuintje het gemiddelde en de variantie van de temperatuurmetingen berekend. Vervolgens worden hypothesetesten uitgevoerd om na te gaan of er correlatie is tussen x (gemiddelde en variantie van de temperatuur) en y (gemiddelde activiteit-densiteit van de arthropodagroepen).
15
5. RESULTATEN
Invloed van de landschapssamenstelling op in de grond
kruipende arthropoda
5.1.1. PCA-analyse
Na uitvoeren van de Bartlett’s test of sphericity kan besloten worden dat de correlaties voldoende groot zijn en een PCA bijgevolg gerechtvaardigd is (p < 0,001). Op de scree plot is er een knik te zien bij de tweede component (Figuur 12, bijlage 9.2). Er wordt bijgevolg besloten om twee componenten te behouden, die samen 55,4% van de totale variabiliteit verklaren. Op figuur 8 is de loading plot te zien die wordt bekomen.
a) b) c)
Figuur 8: Loading plot van de arthropodagroepen met componenten 1 en 2 die samen 55,4% van de totale variantie, verklaren, waarbij component 1 38,2% verklaart en component 2 17,2%. Elke arthropodagroep wordt geplot met als coördinaten de loadings op de eerste twee principale componenten.
Figuur 9: Scatterplot van de scores voor component 1 en component 2 waarbij de datapunten van de tuintjes ingedeeld worden volgens landschapssamenstelling op a) 10, b) 50 en c) 500 meter. De tuintjes werden opgedeeld in HG (hoog groen), LB (landbouw), LG (laag groen) of NG (niet groen) volgens het grootste aandeel.
16
5.1.2. Absence-presence
Er zijn geen opvallende verschillen zichtbaar tussen de boxplots met absence (blauw) en presence (oranje) data (Fig. 10). De hypothesen van de Mann-Whitney U test zijn:
H0: De medianen van absence en presence data zijn gelijk.
H1: De medianen van absence en presence data verschillen van elkaar.
Voorwaarden:
1. Men beschikt over twee onafhankelijke steekproeven van twee populaties. 2. De verdelingen van beide populaties hebben dezelfde vorm.
Tabel 2: p-waarden van de Mann-Whitney U testen. De landschapsvariabelen werden opgesplitst in absence en presence groepen bij miljoenpoten en duizendpoten.
Miljoenpoten Duizendpoten
Percentage niet groen 50 meter 0,50 0,10
Percentage landbouw 50 meter 0,83 0,51
Percentage laag groen 50 meter 0,14 0,27
Percentage hoog groen 50 meter 0,49 0,90
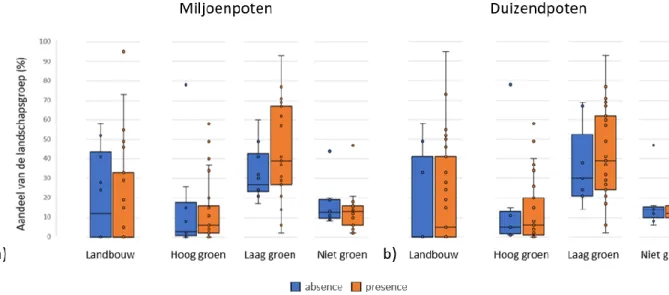
Besluit: Voor alle landschapsvariabelen zijn de p-waarden groter dan 0,05 bij miljoenpoten en duizendpoten (Tabel 2). Op basis van het 5% significantieniveau wordt de nulhypothese aanvaard. Aanwezigheid van miljoenpoten en duizendpoten wordt dus niet bepaald door het omringende landschap op 50 meter.
Figuur 10: Aandeel van de vier landschapsgroepen (landbouw, hoog groen, laag groen en niet groen) op 50 meter bij absence (blauw) en presence (oranje) van a) miljoenpoten en b) duizendpoten. De vier landschapsgroepen werden elk opgedeeld in twee subgroepen die werden bepaald door absence en presence van a) miljoenpoten en b) duizendpoten. Vervolgens werd de verdeling van deze subgroepen gevisualiseerd aan de hand van boxplots bij de vier landschapsgroepen.
17
5.1.3. Meervoudige lineaire regressie
Eerst worden er hypothesetesten uitgevoerd om na te gaan of er regressie is tussen x (het aandeel van de landschapsvariabele) en y (getransformeerde gemiddelde activiteit-densiteit). De nul- en alternatieve hypothesen hierbij zijn:
H0: ρx,y = 0, er is geen correlatie
H1: ρx,y ≠ 0, er is correlatie
Tabel 3: Correlatiecoëfficiënten van de getransformeerde activiteit-densiteit van de arthropodagroepen en landschapsvariabelen op 10, 50 en 500 meter. Significante correlatiecoëfficiënten worden met een * weergegeven.
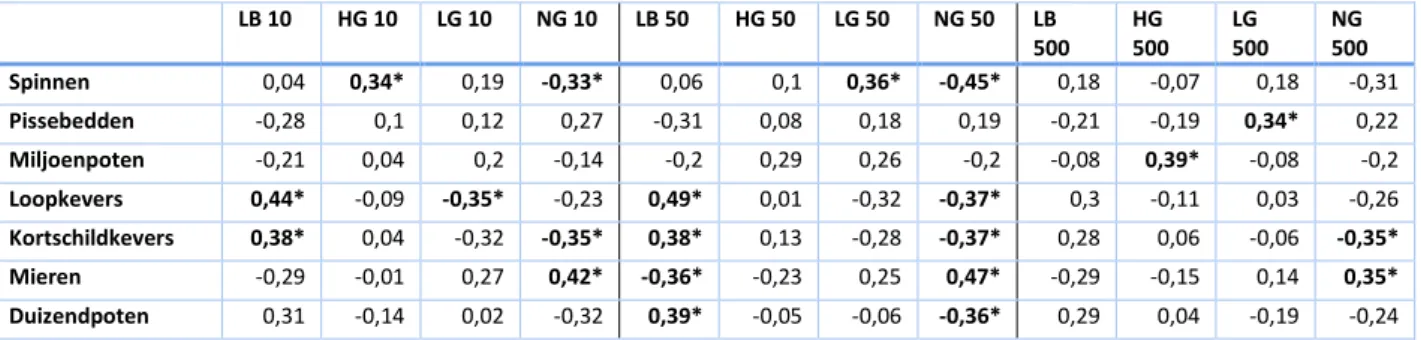
LB 10 HG 10 LG 10 NG 10 LB 50 HG 50 LG 50 NG 50 LB 500 HG 500 LG 500 NG 500 Spinnen 0,04 0,34* 0,19 -0,33* 0,06 0,1 0,36* -0,45* 0,18 -0,07 0,18 -0,31 Pissebedden -0,28 0,1 0,12 0,27 -0,31 0,08 0,18 0,19 -0,21 -0,19 0,34* 0,22 Miljoenpoten -0,21 0,04 0,2 -0,14 -0,2 0,29 0,26 -0,2 -0,08 0,39* -0,08 -0,2 Loopkevers 0,44* -0,09 -0,35* -0,23 0,49* 0,01 -0,32 -0,37* 0,3 -0,11 0,03 -0,26 Kortschildkevers 0,38* 0,04 -0,32 -0,35* 0,38* 0,13 -0,28 -0,37* 0,28 0,06 -0,06 -0,35* Mieren -0,29 -0,01 0,27 0,42* -0,36* -0,23 0,25 0,47* -0,29 -0,15 0,14 0,35* Duizendpoten 0,31 -0,14 0,02 -0,32 0,39* -0,05 -0,06 -0,36* 0,29 0,04 -0,19 -0,24
De hypothesen van de F-test zijn:
H0: De uitbreiding van het model met één predictor verbetert de R2-waarde van het model niet.
H1: De uitbreiding van het model met één predictor verbetert de R2-waarde van het model.
Tabel 4: p-waarden van de F-test weergegeven voor de vier modellen van alle arthropodagroepen. De p-waarden werden afgerond naar 2 cijfers na de komma en significante p-waarden zijn met een * gemarkeerd.
Tabel 5: Regressiemodellen van de zeven arthropodagroepen met de landschapsvariabelen binnen een straal van 50 m. Voor ieder model worden de R2-waarde, de schatter voor het intercept (B0) en de predictoren in volgorde van
toevoeging met hun ongestandaardiseerde coëfficiënt (B) weergegeven. Significante coëfficiënten worden met een * gemarkeerd (p < 0,05). Spinnen B | Predictor Pissebedden B | Predictor Miljoenpoten B | Predictor Loopkevers B | Predictor Kortschild-kevers B | Predictor Mieren B | Predictor Duizendpoten B | Predictor R2 0,20 / 0,19 0,24 0,14 0,33 0,15 B0 2,46* / 0,16 0,84* 0,85* 1,50* 0,30* B1 -0,02* NG / LB 0,01* HG 0,01* LB 0,01* LB 0,03* NG 0,01* LB B2 / LG / NG 0,01* LG / HG / HG 0,01* LG / NG B3 / HG / LG / LB / NG / NG / HG / LG B4 / LB / HG / NG / LG / LG / LB / HG
Spinnen Pissebedden Miljoenpoten Loopkevers Kortschildkev ers Mieren Duizendpote n 1 predictor 0,01* 0,06 0,08 0,00* 0,02* 0,00* 0,02* 2 predictoren 0,05 0,72 0,05* 0,22 0,09 0,03* 0,17 3 predictoren 0,47 0,72 0,46 0,42 0,33 0,44 0,64 4 predictoren 0,14 0,76 0,83 0,45 0,42 0,78 0,56
18 Nagaan van de assumpties voor meervoudige lineaire regressie:
1. Lineariteit van het regressieverband. 2. Onafhankelijkheid van de residuen.
3. Homoscedasticiteit: de residuen hebben een constante variantie. 4. De residuen zijn normaal verdeeld.
5. Er is geen multicollineariteit, de onafhankelijke variabelen zijn niet sterk met elkaar gecorreleerd. In bijlage worden scatterplots voorgesteld van de voorspelde waarden en de residuen (Bijlage 9.3). Er zijn geen duidelijke patronen zichtbaar op deze plots en bijgevolg zijn voorwaarden 1, 2 en 3 voldaan (Figuur 13, bijlage 9.3). Na uitvoeren van de Shapiro-Wilk en de Kolmogorov-Smirnov test, wordt aangetoond dat de residuen normaal verdeeld zijn bij alle regressiemodellen (Tabel 8, bijlage 9.3). Bij de modellen van miljoenpoten en mieren komen er 2 predictor-variabelen voor. Bij miljoenpoten zijn deze variabelen hoog groen en laag groen, bij mieren niet groen en laag groen. Deze predictoren zijn niet significant gecorreleerd met elkaar (p-waarden zijn respectievelijk 0,29 en 0,37) en bijgevolg is de vijfde assumptie ook voldaan. De regressiemodellen van miljoenpoten en mieren bestaan uit meer dan één predictor (Tabel 5). Om te onderzoeken of de interactieterm van deze predictoren significant is, wordt deze toegevoegd aan beide regressiemodellen. De coëfficiënten van de interactietermen bij de regressiemodellen van miljoenpoten en mieren zijn echter niet significant verschillend van 0 (p-waarden zijn respectievelijk 0,61 en 0,97).
Microklimaat
5.2.1. Verband tussen het microklimaat en de landschapssamenstelling
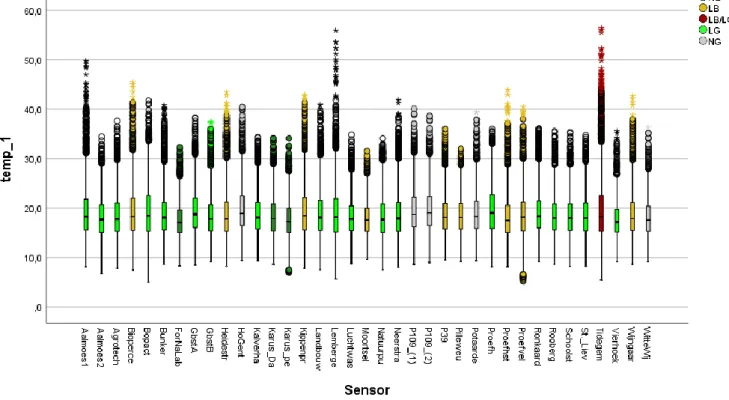
Figuur 11: Boxplots van alle temperatuursmetingen uit 2019 weergegeven per tuintje. Voor de indeling volgens landschap werd de landschapscategorie bepaald met het grootste aandeel op 10, 50 en 500 meter voor alle tuintjes. De finale landschapsgroep voor een tuintje is deze die 2 of 3 keer het grootste aandeel had. Wanneer een tuintje 3 verschillende landschapsgroepen heeft bij 10, 50 en 500 meter, worden deze allemaal weergegeven. LG (lichtgroen): laag groen tuintje, HG (donkergroen): hoog groen tuintje, LB (geel): landbouw tuintje, NG (grijs): niet groen tuintje, rood: ander.
19
5.2.2. Verband tussen arthropoda en microklimaat
De nul- en alternatieve hypothesen hierbij zijn: H0: ρx,y = 0, er is geen correlatie
H1: ρx,y ≠ 0, er is correlatie
Tabel 6: Correlatiecoëfficiënten van activiteit-densiteit van de arthropodagroepen met het gemiddelde en de variantie van de temperatuur. Correlatiecoëfficiënten die significant verschillend zijn van 0 werden gemarkeerd met een *.
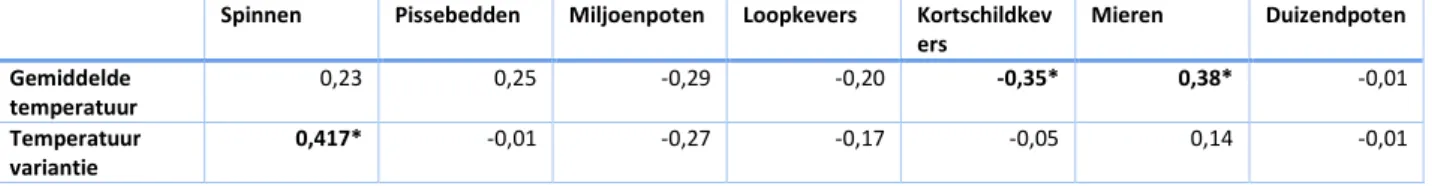
Spinnen Pissebedden Miljoenpoten Loopkevers Kortschildkev ers Mieren Duizendpoten Gemiddelde temperatuur 0,23 0,25 -0,29 -0,20 -0,35* 0,38* -0,01 Temperatuur variantie 0,417* -0,01 -0,27 -0,17 -0,05 0,14 -0,01
20
6. DISCUSSIE
Wat is het effect van het percentage landbouw/niet groen/laag
groen/hoog groen op arthropoda?
Bij het uitvoeren van de PCA-analyse werden twee componenten behouden. Op de loading plot (Figuur 8) is te zien dat de eerste component voornamelijk bepaald wordt door de activiteit-densiteit van mieren aan de negatieve zijde en de activiteit-densiteit van loopkevers, kortschildkevers en duizendpoten aan de positieve zijde van de as. Op de scatterplots (Figuur 9) is te zien dat tuintjes met hoofdzakelijk niet-groen, hoog groen en landbouw zich voornamelijk positioneren respectievelijk aan de negatieve zijde, in het midden en aan de positieve zijde van de eerste as. Wellicht zal de activiteit-densiteit van mieren laag zijn in tuintjes met veel landbouw, en deze van loopkevers, kortschildkevers en duizendpoten hoog. Het omgekeerde verschijnsel zal zich voordoen in tuintjes met veel niet groen. De tweede component wordt voornamelijk bepaald door de activiteit-densiteit van pissebedden (Figuur 8). Omdat de tuintjes met veel landbouw zich verzamelen aan de onderzijde van de scatterplots (Figuur 9), zal de activiteit-densiteit van pissebedden waarschijnlijk laag zijn in deze tuintjes. Deze vermoedens uit de exploratieve PCA-analyse worden verder geverifieerd en besproken aan de hand van de regressiemodellen.
We verwachten dat arthropodagroepen met soortgelijke functies gelijkaardig zullen reageren op omgevingsinvloeden. In de onderordes zullen de arthropodagroepen aldus worden besproken volgens ecologisch functionele groep. Loopkevers, kortschildkevers en duizendpoten gedragen zich gelijkaardig op de loading plot en worden samen besproken bij de onderorde ‘predatoren’. Ook spinnen zijn predatoren en worden in deze sectie besproken. Miljoenpoten en pissebedden worden besproken bij de onderorde ‘detritivoren’. Mieren zijn zonderlinge arthropoda op de loading plot en worden apart besproken.
6.1.1. Predatoren
Loopkevers, kortschildkevers en duizendpoten zijn nauwelijks gecorreleerd met het percentage hoog groen, negatief met het percentage laag groen en niet groen, en positief met het percentage landbouw (Tabel 3). Landbouwpercelen die intensieve monoculturen zijn, en weinig onder druk staan van agrochemicaliën, herbergen tijdens het groeiseizoen gewoonlijk dense populaties plagen en natuurlijke vijanden (Altieri & Nicholls, 2003). Hoewel we niet over informatie beschikken met betrekking tot gebruik van agrochemicaliën, wordt in onze resultaten ook een positieve invloed teruggevonden van het landbouwpercentage op de activiteit-densiteit van deze predatoren (Tabel 3,
Tabel 5). Na de oogst en voor het nieuwe groeiseizoen zijn landbouwpercelen ongeschikt als habitat. Volgens Duflot et al. (2016) is de aanwezigheid van complementaire, permanente landschapselementen van waaruit migratie mogelijk is dan ook bepalend voor de populatiegrootte van predatoren. Dit kan onder de vorm van hoog groene elementen zoals hagen en houtkanten. Voor de predatoren zijn dit waardevolle, permanente landschapselementen waar alternatieve voedselbronnen en schuilplaatsen beschikbaar zijn (Tscharntke et al., 2005; Bianchi et al., 2006). Het positieve effect van het percentage hoog groen op de activiteit-densiteit van loopkevers, kortschildkevers en duizendpoten is in onze resultaten echter nauwelijks zichtbaar (Tabel 2). In de concentrische cirkels van 50 meter was bij ieder tuintje steeds minstens 7% semi-natuurlijke habitat (laag groen en hoog groen) aanwezig. Mogelijks volstaat een kleine hoeveelheid hoog groen als habitat, en een overheersende hoeveelheid hoog groen reduceert de proportie landbouw in het landschap. Ook laag groene elementen zoals grasstroken hebben een belangrijke functie in het agro-ecosysteem (Marshall & Moonen, 2002) en kunnen als habitat gebruikt worden door de predatoren (Holland et al., 2009). Anderzijds zouden natuurlijke weilanden en private tuinen, die ook onder de laag groene elementen geklasseerd worden, weinig geschikte habitats zijn (Caro et al., 2016). Vooral het maaien van deze laag groene elementen veroorzaakt sterke habitatdestructie, met nadelige gevolgen voor de activiteit-densiteit van arthropoda
21 (Dennis et al., 2015). Deze bevindingen kunnen verklaren waarom de activiteit-densiteit van loopkevers en kortschildkevers negatief gecorreleerd is met het percentage laag groen op 10 en 50 meter (Tabel 3). Spinnen zijn ook predatoren, maar hun gedrag is anders dan dat van de eerder besproken predatoren (Figuur 8). In het regressiemodel van spinnen is te zien dat de activiteit-densiteit voornamelijk bevoordeeld wordt door de afwezigheid van niet groen (
Tabel 5). Spinnen zijn zeer gevoelig aan vervuiling en staan bekend voor hun rol als bio-indicatoren (Marc et al., 1999). Het is dus niet merkwaardig dat ze minder teruggevonden werden in tuintjes met veel niet-groen in de omgeving. Spinnen zijn zeer waardevolle natuurlijke vijanden in een landbouwgebied. Zowel de activiteit-densiteit als de soortenrijkdom is groot in agro-ecosystemen waardoor ze een heterogene predatorgroep vormen met sterk uiteenlopende jachtstrategieën, habitatvoorkeuren en actieve periodes (Marc et al., 1999; Bhomwick & Abrol, 2017). Dankzij de ‘ballooning’-eigenschap (waarbij ze zich passief laten meevoeren met de wind) zijn ze heel mobiel en horen ze bij de eerste arthropoda om een verstoord landbouwperceel opnieuw te koloniseren (Bishop & Riechert, 1990; Öberg & Ekbom, 2006). In onze resultaten werd echter geen significant positieve correlatie teruggevonden tussen het landbouwpercentage en de activiteit-densiteit van spinnen, wat ook aantoont dat ze verschillen van de andere predatoren (Tabel 3). Volgens Wise (1995) hebben spinnen een divers dieet nodig en wordt hun populatiegrootte eerder bepaald door de beschikbare prooidiversiteit dan door de populatiegrootte van prooien. Dit verklaart waarom de activiteit-densiteit van spinnen weinig bepaald werd door de proportie gesimplificeerd landbouwlandschap met weinig niches en aldus een lage (prooien) soortendiversiteit (Mestre et al., 2018), maar veeleer door de percentages laag groen en hoog groen (Tabel 3). De positieve correlatie van hoog groen met de activiteit-densiteit van spinnen is voornamelijk merkbaar op kleine schaal (Tabel 3) omdat het de structuurrijkdom zou verhogen, en er aldus hoge prooiendiversiteit beschikbaar zou zijn voor de spinnen binnen jachtbereik (Marc et al., 1999).
6.1.2. Detritivoren
Een studie van Vilisics et al. (2007) over het effect van urbanisatie op de pissebeddenpopulaties in Sorø toonde aan dat verstedelijkte gebieden een geschikte habitat kunnen vormen voor pissebedden. Sommige pissebeddensoorten hebben een specifieke voorkeur voor droge en verstoorde gebieden. Dit zou zijn omwille van de grote diversiteit aan microhabitat en grote hoeveelheid kwaliteitsvolle voedselbronnen in verstoorde gebieden (Rushton & Hassall, 1983). Andere soorten kwamen in gelijke hoeveelheden voor op de
verschillende locaties, dit toont aan dat bij sommige pissebeddensoorten geen specifieke habitatvoorkeur aanwezig is (Vilisics et al., 2007). Uit de PCA blijkt dat de activiteit-densiteit van pissebedden laag zal zijn in tuintjes met veel landbouw, maar in de regressieanalyse wordt dit effect niet significant bevonden (Tabel 3, Tabel 5). Bodemverstoring is frequent aanwezig in landbouwpercelen en heeft nadelige gevolgen voor pissebeddenpopulaties (Souty-Grosset & Faberi, 2018). Anderzijds was de gewone oprolpissebed (Armadillidium vulgare) de dominante soort in onze bodemvallen, een droogtetolerante soort die algemeen voorkomt in landbouwlandschap (Faberi et al., 2011). Een andere soort die veel aanwezig was in de bodemvallen was de ruwe pissebed (Porcellio scaber), dit is een soort die weinig voorkomt in landbouwpercelen (Souty-Grosset & Faberi, 2018). De diversiteit aan habitatvoorkeuren binnen de groep van de pissebedden kan verklaren waarom onze resultaten geen significante invloed van het landschap op de activiteit-densiteit van pissebedden aantonen. Het is ook mogelijk dat de activiteit-densiteit veeleer werd bepaald door het microklimaat (Meinertz, 1950; Gruner, 1966) of de pH en de microbiële activiteit in de strooisellaag (Zimmer & Topp, 1997).
Miljoenpoten komen voornamelijk voor in de strooisellaag van loofbossen, het zijn detritivoren, saprofyten of fytofagen met het strooisel als voornaamste voedselbron (Kime & Golovatch, 2000). Hornak et al. (2020) toonden aan dat de dikte van de strooisellaag een significant effect heeft op de activiteit-densiteit van miljoenpoten. In de resultaten van dit onderzoek is het dan ook niet verrassend dat er een positieve correlatie teruggevonden wordt tussen de activiteit-densiteit van miljoenpoten en het percentage hoog groen.