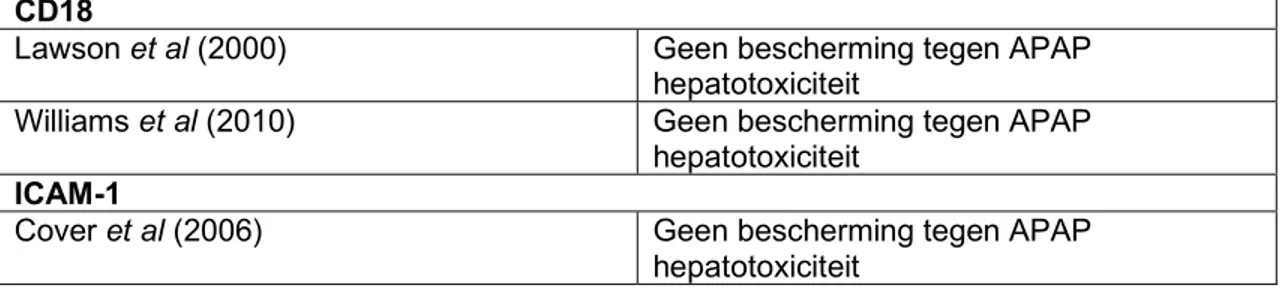
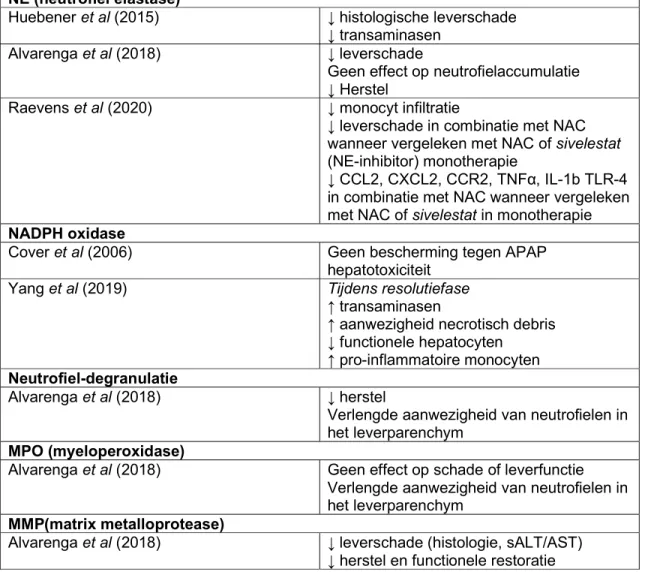
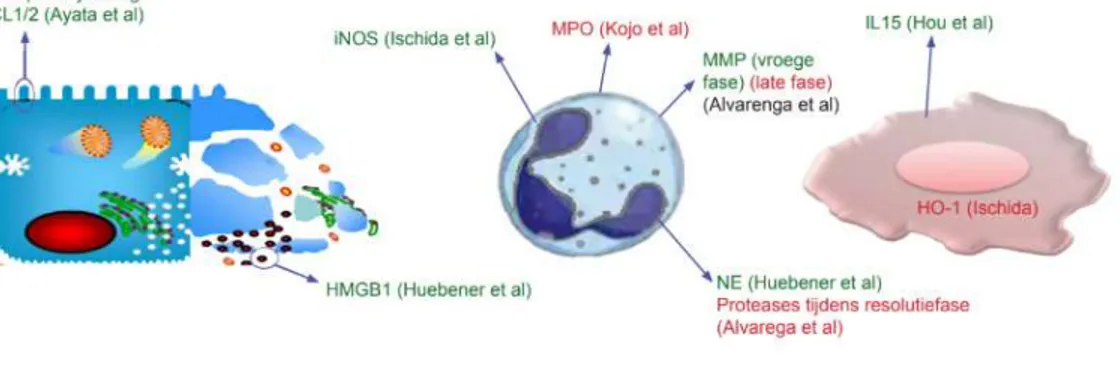
De neutrofiel in acute paracetamol
geïnduceerde leverziekte
Lievens Louis
Stamnummer: 01508519Promotor: Prof. dr. Hans Van Vlierberghe Copromotor: dr. Sarah Raevens
Masterproef voorgelegd in het kader tot het behalen van de graad Master of Medicine in de Geneeskunde
1
Inhoudsopgave
Abstract ... 2
Inleiding ... 4
Acute leverziekte ... 4
1) Acute leverziekte en acuut leverfalen (ALF) ... 4
2) Epidemiologie ... 4
3) Etiologie ... 5
4) Pathogenese van acute leverziekte ... 6
5) De pathogenese van acute acetaminophen-geinduceerde leverziekte ... 11
6) Acute leverziekte: huidige aanpak en therapiemogelijkheden ... 13
De neutrofiel in acute leverziekte ... 15
1) De neutrofiel in het immuunsysteem ... 15
2) De neutrofiel in de lever ... 15
a) Rekrutering naar de levervasculatuur ... 15
b) Extravasatie naar het leverparenchym ... 16
c) Werking van neutrofielen in de lever ... 17
d) De neutrofiel en resolutie van inflammatie ... 19
Vraagstelling ... 20
Methodologie ... 21
Resultaten ... 22
1) Argumenten voor een rol van neutrofielen in het exacerberen van APAP geïnduceerde leverschade ... 22
2) Argumenten tegen een rol voor neutrofielen in de exacerbatie van APAP-geinduceerde leverschade ... 32
3) Een dynamische rol van neutrofielen in APAP-hepatotoxiciteit, onderhevig aan het inflammatoire milieu waarin zij zich bevinden ... 40
Discussie ... 44
2
Abstract
Acuut leverfalen (ALF) is de klinische uiting van ernstige leverschade, waarbij het verlies van immunologische en metabole functie levensbedreigende gevolgen kan hebben. Paracetamol is in de westerse wereld de meest voorkomende oorzaak van ALF, waarbij de huidige therapeutische opties eerder beperkt zijn, en een urgente levertransplantatie (ELT; emergency liver transplantation) soms als laatste redmiddel ingeschakeld moet worden.
Steriele inflammatie en necrose staan centraal in de pathogenese van paracetamol-geïnduceerd leverfalen, waarbij de vrijstelling van damage associated molecular patterns (DAMPs) het immuunsysteem activeert tot de productie van inflammatoire cytokines en chemokines leidend tot de rekrutering van immuuncellen, die potentieel de schade kunnen exacerberen of regeneratie stimuleren.
De neutrofiel hoort bij de eerste groep cellen die naar de inflammatoire zone gerekruteerd wordt, en deze groep heeft het vermogen om schade te verergeren via onder andere adherentie-dependente cytotoxiciteit via de vorming van reactive oxygen species (ROS) of degranulatie, maar heeft mede een regulerende rol in het proces van de inflammatie, door andere immuuncellen aan te sturen. Een belangrijke rol voor de neutrofiel in het verwijderen van debris en het reguleren van resolutie en regeneratie is eerder aangehaald.
Het doel van deze masterproef is om via een literatuurstudie te onderzoeken of interventies tegen de neutrofiel kunnen leiden tot het boeken van therapeutische winst bij paracetamol-geïnduceerd leverfalen.
Resultaten: Meerdere studies onderzochten het inflammatoire proces rond paracetamol-geïnduceerd leverfalen. Verschillende interventies tegen DAMPs, interleukines, chemokines, pattern recognition receptors (PRRs) en adhesiemoleculen betrokken bij neutrofielfunctie werden gecorreleerd met leverschade en neutrofielinfiltratie, pleitend voor een exacerberende rol van deze cellen in de pathogenese. Echter, andere studies publiceerden vaak tegenstrijdige resultaten, waarbij interventies tegen bepaalde DAMPs, interleukines, chemokines, PRRs en andere geen bescherming boden tegen paracetamol-geïnduceerde hepatotoxiciteit, en waaruit niet besloten kon worden dat neutrofielen significant bijdroegen tot de ziekteprogressie. Verder werden verschillende fysiologische mechanismen aangehaald die potentiele schade-exacerbatie door neutrofielen verhinderden, en ook een beschermende rol voor neutrofielen werd meermaals geïllustreerd, met belangrijke functies in regeneratie, resolutie en de regulatie van andere immuuncellen. Interventies tegen effectorfuncties van de neutrofiel leidden vaak tot een verminderde inflammatie en leverschade, maar vaak werd gezien dat dit ook negatieve gevolgen had voor regeneratie en resolutie. Andere studies
3 toonden aan dat het gedrag van de neutrofiel sterk variabel is naargelang het inflammatoire milieu en de aanwezigheid van andere regulerende immuuncellen.
Discussie en besluit: Er is voldoende evidentie om te stellen dat een relatie bestaat tussen de aanwezigheid van inflammatoire merkers, de infiltratie van neutrofielen in het inflammatoir milieu en het optreden van leverschade, waaruit vermoed kan worden dat steriele inflammatie wel degelijk een rol speelt in de progressie van leverschade – en falen. Wel kan uit deze masterproef niet besloten worden dat neutrofielen in dit proces oorzakelijk zijn, en moet gesteld worden dat neutrofielen inderdaad bijdragen tot resolutie, regeneratie en modulatie van het inflammatoire proces. Valkuilen in verschillende studies zijn verschillen in opstel van experimenten (type muis, type cel, dosis paracetamol, wijze van toediening), het niet evalueren van eventuele negatieve effecten van interventies tijdens resolutie en regeneratie, alsook het niet evalueren van de effecten van de interventies op het metabolisme van paracetamol in de hepatocyt, waardoor interpretatiefouten kunnen ontstaan. Ook werd vaak nagelaten om de exacte bijdrage van neutrofielen in het proces te onderzoeken, waardoor vaak enkel een correlatie werd aangetoond zonder dat deze mechanistisch verklaard wordt. In conclusie is er een duidelijke associatie tussen inflammatie en de progressie van paracetamol-geïnduceerde leverschade. De rol van neutrofielen in dit proces kan niet eenduidig gesteld worden, en is waarschijnlijk afhankelijk van het inflammatoire milieu waarbinnen de neutrofiel zich dichotoom kan gedragen. Interventies tegen neutrofielen hebben het potentieel om therapeutische winst te boeken in de pathogenese, al moet steeds rekening gehouden worden met eventuele effecten op resolutie en regeneratie, alsook een verhoogde vatbaarheid voor secundaire infecties bij ALF.
4
Inleiding
Acute leverziekte
1) Acute leverziekte en acuut leverfalen (ALF)
Acute leverziekte kan ontstaan uit diverse ethologiën, en kan bij een ernstig verloop een beeld van acuut leverfalen veroorzaken (ALF; acute liver failure). ALF is de klinische uiting van ernstige leverschade, wat door het verlies aan metabole en immunologische functie levensbedreigende gevolgen kan hebben. De kliniek omvat een verlengde prothrombinetijd/international normalized ratio (PT/INR) met coagulopathie, portale hypertensie, neurologische symptomen door encefalopathie met cerebraal oedeem en intracraniële hypertensie, perifere vasodilatatie en hypotensie, immuundysfunctie leidend tot een verhoogd risico op sepsis, SIRS (systemic inflammatory response syndrome) en multipel orgaanfalen (MOF) met een risico op overlijden van de patiënt. Vanwege een verlies aan metabole leverfunctie wordt ook vaak hypoglycemie, lactaatacidose en hyperammoniemie gezien. (1-3)
Voor ALF zijn zeer veel definities en criteria ontwikkeld gedurende de jaren, wat in de klinische praktijk gebruikt wordt om de prognose en dus de nood tot transplantatie in te schatten. In de meeste classificatiesystemen staat het optreden van hepatische encefalopathie (HE) na het begin van de symptomen (vaak het optreden van geelzucht) centraal. (4)
2) Epidemiologie
Acute leverziekte en ALF zijn zeldzame aandoeningen in de ontwikkelde wereld, omdat de infectieuze oorzaken dankzij specifieke gezondheidsmaatregelen (sanitair, vaccinaties) in de Westerse wereld een minder grote impact vertonen. Dit wordt geïllustreerd door wereldwijde geografische variaties: in Europa en de Verenigde Staten is er een hoge incidentie van medicijn-geïnduceerd leverfalen, terwijl in de ontwikkelingslanden de incidentie van de virale hepatitisvirussen (voornamelijk HAV, HEV en HBV) hoger blijft. (1, 5) Wereldwijd blijven de virale hepatitisvirussen de grootste oorzaak van ALF. (6) De meer zeldzame ethologiën zijn in 11%-23% de oorzaak van ALF, afhankelijk van de definitie van ‘zeldzaam’. (4) De incidentie van de verschillende oorzaken evolueren met de tijd omdat ze afhankelijk zijn van bevolkingsvaccinatie, sanitaire voorzieningen, de beschikbaarheid van en de regelgeving omtrent farmacologische producten.
5 Tabel 1: Epidemiologie van ALF in verschillende landen. [Overgenomen uit (5)]
3) Etiologie
Zoals aangehaald zijn er verschillende oorzaken mogelijk, die wereldwijd geografisch variëren. Virale hepatitis (voornamelijk HAV en HBV), medicijn-geïnduceerd leverfalen, idiosyncratische reacties op medicijnen, toxines en metabole oorzaken zijn de meest voorkomende oorzaken wereldwijd. (7) Van de medicijn-geïnduceerde acute leverziekte is acetaminophen (paracetamol) de meest voorkomende in de Verenigde Staten en Noord Europa, waarbij de leverschade dosis-afhankelijk is. (2, 3)
Na paracetamol zijn de andere vormen van medicijn geïnduceerd leverfalen geografisch verschillend, maar vaak worden antibiotica, ecstasy, tuberculostatica, niet steroïdale anti-inflammatoire middelen (NSAID`s) en aminosalicylaten als oorzaak gevonden. (7) Ook voedings- en kruidensupplementen, alsook chemotherapeutische, immunosuppressieve en antiretrovirale medicatie worden als potentieel hepatotoxisch beschouwd. (8)
Figuur 1: Wereldwijde oorzaken van ALF [overgenomen uit (3)]
6 Niet enkel de klassieke hepatitis virussen (HAV, HBV, HCV, HEV) kunnen ALF veroorzaken, maar ook herpes simplex virus 1 en 2 (HSV 1 en 2), humaan herpes virus 6, het varicella zoster virus, het Ebstein Barr virus (EBV), het cytomegalievirus (CMV) en parvovirus B16 kunnen dit veroorzaken. (2)
Andere (meer zeldzame) oorzaken van ALF zijn hyperthermie na hitteshock, specifieke toxines (bv paddenstoel), metabole aandoeningen (bv ziekte van Wilson), auto-immuun hepatitis, ischemische schade bij systemische hypotensie (na sepsis, hartfalen, chronische respiratoire decompensatie) of het Budd Chiari syndroom. Ook maligne infiltratie van de lever, het HELPP syndroom (hemolysis syndrome, elevated liver enzymes and low platelets) bij zwangerschap en het Reye syndroom kunnen ALF uitlokken. (2)
Alcoholische hepatitis (AH) is een vorm van acute leverinflammatie die kan voorkomen bij patiënten die een grote hoeveelheid alcohol innemen, en valt onder het spectrum van de alcoholische leverziekten, waartoe leververvetting (hepatosteatose), alcoholische hepatitis en cirrose behoren. (9)
4) Pathogenese van acute leverziekte
a) Inflammatie: activatie van het immuunsysteem door DAMPs, PAMPs en het inflammasoom
De etiologie is bepalend voor hoe het immuunsysteem reageert op acute leverziekte. Afhankelijk van de originele schade, gaan PAMPs (pathogen associated molecular patterns) of DAMPs (damage associated molecular patterns) een immuunreactie starten. (10)
In de afwezigheid van pathogenen spreekt men over steriele ontsteking (sterile inflammation, SI), zoals dat in de lever gezien wordt bij medicijn geïnduceerd leverfalen, (non-)alcoholische steatohepatitis, fibrose en carcinogenese. DAMPs worden vrijgesteld bij leverschade en activeren na binding op receptoren de immuunreactie via de productie van cytokines en de rekrutering van immuun cellen naar de ontstekingshaard. Bijvoorbeeld, molecules afkomstig van de mitochondriën (uitgebreid aanwezig in de metabool actieve lever) zoals ATP, mitochondriaal DNA, cytochroom c en formylpeptide zijn gekende DAMPs. Het is aangetoond dat verschillende cytosolische eiwitten (waaronder DAMPs en cytokines) onderhevig zijn aan de redox-status van het cytosol, waardoor deze in normale omstandigheden inactiever zijn als immunomodulerende mediatoren, en in een minder gunstige (necrotische) omgeving de inflammatie kunnen sturen en verlengen. (11) Bijvoorbeeld, HMGB1 kan afhankelijk van diens redox-status werken als chemokine, of net werken als pro-inflammatoir cytokine. (12) Tissue factor (TF) kan niet alleen geproduceerd worden voor neutrofielen, maar kan deze ook activeren. (13) Ook complement wordt geactiveerd door de vrijstelling van DAMPs. Zowel cytokines als complementfactoren (waaronder C5a) zijn potente activators voor allerhande immuuncellen, zoals bijvoorbeeld Kupffer cellen en neutrofielen, die geactiveerd worden om
7 ROS (reactive oxygen species; zie later) te produceren en via opregulatie van adhesiemolecules de rekrutering van meerdere immuuncellen naar de lever vasculatuur starten. (14) PAI-1 (plasminogeen activator inhibitor 1) heeft niet enkel een inhiberende werking op fibrinolyse, maar kan ook neutrofielmigratie induceren en reguleren in bepaalde omstandigheden. (15)
Zowel de DAMPs als de PAMPs (specifieke herkenningspatronen van mico-organismen, bijvoorbeeld lipopolysacharide (LPS)) activeren en moduleren de immuunreactie via hun respectievelijke receptoren, de PRRs (pattern recognition receptor). Een goed bestudeerde groep van PRRs zijn de TLRs (toll like receptoren), die zowel DAMPs als PAMPs kunnen herkennen, waarbij bijvoorbeeld TLR9 zich intracellulair bevindt en een hoge affiniteit heeft voor oligodeoxynucleotiden (kenmerk van bacterieel DNA), terwijl TLR4 en TLR2 eerder voorkomen op het celoppervlak om bijvoorbeeld LPS te herkennen. (14, 16, 17)
Verschillende cellen uit de lever zijn in staat necrotische DAMP en PAMP-signalen te herkennen en een inflammatie op gang te brengen. Het is aangetoond in verschillende modellen dat lever-residente myeloïde levercellen (Kupffer cellen, macrofagen, dendrietische cellen), vasculaire endotheelcellen en stromale (steun)cellen PRRs tot expressie brengen om gevaarsignalen te herkennen, en zo via de productie van cytokines de inflammatie op gang kunnen brengen. (11) Na de activatie door de eerste stimulus (PAMPs, DAMPs) worden effectorcellen van het aangeboren immuunsysteem (monocyten, macrofagen en neutrofielen) aangetrokken tot de infectiehaard, waar deze cellen de inflammatie onderhouden via de productie van ROS en pro- en anti-inflammatoire cytokines. (10)
Een belangrijke bedenking is het feit dat het aangeboren immuunsysteem niet als doel heeft de originele schade te verergeren, maar eerder een opruimfunctie heeft, zoals het verwijderen van dode en stervende cellen. (14)
Het inflammasoom is een cytosolisch multi-eiwit complex dat belangrijk is voor de activatie van caspase-1 en de initiatie van een ontstekingsreactie via de productie van inflammatoire interleukines. Om geactiveerd te kunnen worden zijn 2 signalen vereist, waarvan het eerste de transcriptionele opregulatie van pro-cytokines en inflammasoom onderdelen via TLRs/PRRs en de NFκβ pathway is. Het tweede signaal kan verschillende triggers hebben, waaronder ATP, ROS en micropartikels zoals urinezuur. De twee signalen samen activeren het caspase-1, dat de pro-inflammatoire cytokines pro-IL1β en pro-IL18 activeert en hun secretie start. (11)
8 Figuur 2: Overzicht van steriele
inflammatie.
[overgenomen uit (10)] A) Celnecrose leidt tot de vrijstelling van DAMPs, waardoor immuuncellen
geactiveerd worden tot de productie van inflammatoire cytokines leidend tot de initiatie van een
ontstekingsreactie, potentieel leidend tot immuun gemedieerde celschade
B) Twee signalen zijn nodig om het
inflammasoom te activeren. Het eerste signaal activeert via onder andere NFκβ de
transcriptie van pro-inflammatoire cytokines (pro-IL-1β en pro-IL-18) en inflammasoom-onderdelen, het tweede signaal activeert het caspase-1. Beide signalen kunnen door verschillende molecules geactiveerd worden. Afkortingen: DAMP = damage associated molecular pattern; TNF = tumor necrosis factor; TNFR = TNF receptor ; TLR = toll like receptor ; ATP = adenosine trifosfaat ; NLRP = NOD-, LRR- and pyrin domain-containing protein ; ASC = apoptosis associated speck like protein; P2X7 = P2X purinoceptor 7
b) Het geactiveerd immuunsysteem: mechanismen van celdood
ALF treedt op wanneer er meer hepatocyten afsterven dan dat er regeneratie optreedt, met verlies aan hepatische functionaliteit. Celdood kan op verschillende manieren optreden, en welke vorm van celdood optreedt (apoptose, overleving, necrose, pyroptose) is waarschijnlijk afhankelijk van de etiologie en de aangesproken signaaltransductiecascades. (10, 18) Apoptose is een gereguleerde celdood die verloopt via een interne proteolytische digestie, geactiveerd door ‘cell death pathways’ die leiden tot de activatie van caspases. Liganden als TRAIL, CD95L (soluble death receptor) of TNFα kunnen deze ‘death pathway’ activeren via receptorbinding. Dit is echter niet absoluut; TNF-α speelt ook een rol in lever regeneratie en inflammatie via de activatie van het NFκβ gen. Hoewel apoptose beschreven wordt als een
9 gereguleerd proces van celdood, kan inadequate en overweldigende activatie van ‘cell death pathways’ een beeld van ALF veroorzaken wegens verlies aan leverfunctie. Bij medicijn-geïnduceerd leverfalen wordt de apoptose voornamelijk medicijn-geïnduceerd via een intrinsieke pathway, die geactiveerd wordt door mitochondriale schade. (18)
Figuur 3: vereenvoudigd schema van celdood in acuut leverfalen.[overgenomen uit (18)]
TNF receptor-1 activatie kan leiden tot NFκβ activatie en overleving, apoptose of necroptose, en het resultaat is afhankelijk van de receptor geassocieerde signaalcomplexen. Bij apoptose wordt het signaal geamplificeerd via de mitochondriën, waar een translocatie van Bax en Bak (geactiveerd door het caspase-8 gemedieerde klieven van Bid) leidt tot permeabilisatie van de buitenste mitochondriale membraan, het loslaten van cytochroom c en caspase-3 activatie. Necroptose kan voorkomen bij een vermoeilijkte apoptose, wat gemedieerd wordt via RIP-1 en RIP-3 kinases, waarbij een toegenomen oxidatieve stress via ROS (reactive oxygen species) productie leidt tot een intracellulaire calcium stijging, het openen van MPT (mitochondrial membrane permeability transition pore), ATP (adenosine trifosfaat) depletie en uiteindelijk necrose. APAP – zie infra.
Necrose staat tegenover apoptose, en wordt gemedieerd door het openen van de MPT (mitochondrial membrane permeability transition pore), wat leidt tot een onderbreking van de membraanpotentiaal en de vorming van ATP (adenosine trifosfaat) verstoort. Hierdoor zwelt het mitochondrium, waardoor dit uiteindelijk gaat ruptureren en gaat leiden tot een vrijstelling van intermembranaire eiwitten en gefragmenteerd genetisch materiaal. Necrose gaat gepaard met een toename in het intracellulair calcium, verminderde ATP vorming en een toegenomen intracellulaire oxidatieve stress: kenmerken die de necrose mee bepalen en potentieel de necrose exacerberen. Belangrijk is dat necrose vaak gepaard gaat met een inflammatiereactie, als reactie op de vrijstelling van intracellulaire celcomponenten (DAMPS), terwijl dit bij apoptose minder prominent is door een gecontroleerde fagocytose van de apoptotische cellen met bijgevolg een verminderde loslating van DAMPs. (18)
Necroptose is een mengvorm tussen apoptose en necrose die kan geïnduceerd worden door specifieke stimuli, zoals TNF. Bij necroptose worden ook DAMPs vrijgesteld, maar het is een proces dat strikt gereguleerd wordt op verschillende niveaus, en het wordt beschouwd als een gecontroleerde inflammatoire vorm van celdood. (11)
Pyroptose is een andere inflammatoire vorm van celdood waarbij cellyse en de loslating van verschillende pro- en anti-inflammatoire cytokines (waaronder IL-1β en IL18) optreedt. Pyroptose kan op twee manieren geactiveerd worden: via inflammasoom-activatie (zie supra)
10 en via caspase-11, dat geactiveerd wordt door interactie met LPS (lipopolysacharide). Deze vorm van celdood doet vooral dienst om geïnfecteerde cellen tijdig te verwijderen, maar net als bij apoptose kan een excessieve pyroptose activatie leiden tot een verergering van de leverschade, wat bij acute leverziekte relevant is wegens een verhoogde leverexpositie aan micro-organismen (en dus LPS), door onder andere een toename van de intestinale permeabiliteit en immuundysfunctie. (19, 20)
c) Inflammatie: schade door oxidatieve stress
Onder bepaalde omstandigheden is aangetoond dat geactiveerde immuuncellen (neutrofielen, Kupffer cellen, monocyten/macrofagen) de oorspronkelijke schade kunnen verergeren. De vorming van ROS (reactive oxygen species) door deze immuuncellen staat centraal in de pathofysiologie van inflammatie, wat via een intracellulaire oxidatieve stress necrose induceert door het veroorzaken van mitochondriale dysfunctie. (14)
ROS kan op verschillende manieren gevormd worden. Elektronen reducties van zuurstof leidt tot de vorming van superoxide (O2-), waterstof peroxide (H2O2) en hydroxylradicalen. Verder
kan ROS ook gevormd worden door het myeloperoxidase, dat waterstof peroxide omzet tot hypochloorzuur (HOCl). LPO (lipid peroxidation) vormt ROS producten zoals bijvoorbeeld vetzuur hydroperoxide. Ook kan peroxynitriet gevormd worden door de combinatie van superoxide en stikstofmonoxide (NO), wat een RNS (reactive nitrogen species) is, en RNS kan ook leiden tot de vorming van nitrotyrosine. Fagocyten bevatten NADPH (nicotinamide adenine dinucleotide fosfatase) oxidase (NOX2), dat bij activatie superoxide in het fagosoom of buiten de cel brengt. ROS en RNS kunnen op vele manieren binnen en buiten de cel gevormd worden. (21)
Cellen hebben verdedigingsmechanismen ontwikkeld tegen de potentieel schade-inducerende ROS en RNS producten. Bijvoorbeeld, superoxide (O2-) kan verwijderd worden door SOD1
(Cu/Zn superoxide dismutase) of SOD2 (Mn superoxide dismutase) uit de mitochondriën, waardoor minder peroxynitriet (ONOO-) gevormd wordt. Een overzicht van de pro-oxidatieve en anti-oxidatieve werking van een cel wordt gegeven in figuur 4. (21) Verdere anti-oxidatieve bescherming in hepatocyten wordt gemedieerd door onder andere het HO-1 (haem oxygenase-1). (22)
11 Figuur 4: cellulaire mechanismen van ROS vorming en anti-oxidatieve verdediging. [overgenomen uit (21)]
Superoxide wordt vooral gevormd uit het elektronentransport van de mitochondriën, wat via SOD 1&2 omgezet kan worden tot waterstof peroxide of peroxynitriet. Vervolgens kan waterstofperoxide omgezet worden tot een hydroxylradicaal, wat vervolgens LPO (vet peroxidatie) kan activeren, of hypochloorzuur vormen via het MPO (myeloperoxidase). Waterstofperoxide kan echter ook geantioxideerd worden via een catalase, het glutathion peroxidase of peroxyredoxine. GSH (glutathion) heeft een anti-oxidatieve werking via het glutathione-peroxidase of via directe binding met peroxynitriet of hypochloorzuur. Vitamine E werkt anti-oxiderend via de onderbreking van LPO vorming. De aanwezigheid van ijzer (Fe2+) stimuleert LPO.
Lever-residente en gerekruteerde immuuncellen kunnen een extracellulaire bron van ROS zijn. Vermits Kupffer cellen (residente macrofagen) gefixeerd in de sinusoïden zitten, wordt door hen geproduceerd ROS vrijgelaten in de vasculatuur en in de ruimte van Disse, waardoor hepatocyten met de loslating van GSH (glutathione) de oxidatieve stress kunnen neutraliseren. Echter, geëxtravaseerde neutrofielen (zie later) adhereren zich aan de doelcel, waardoor zij volledig geactiveerd worden en vervolgens een langdurige oxidatieve stress kunnen veroorzaken en onderhouden, met een rechtstreekse diffusie van ROS in de hepatocyt en het in stand houden van intracellulaire stress. (21)
ROS kan op verschillende manieren celdood induceren. De activatie van apoptotische pathways zoals JNK (c-jun-N-terminal kinase) en caspases door de vorming van superoxide en oxidatieve stress is eerder gedemonstreerd in cellijnen. Echter, in vivo experimenten die gebruik maakten van rattenlevers toonde geen apoptose bij een toegenomen intracellulaire superoxide concentratie; eerst trachtten de cellen via hun verdedigingsmechanisme (bijvoorbeeld GSH) te detoxifiëren met het optreden van necrose wanneer de verdedigingsmechanismen insufficiënt leken te zijn. Necrose uitgelokt door ROS heeft zoals klassieke necrose te maken met MPT opening, verbreking van de mitochondriale membraanpotentiaal en ATP depletie. De gevoeligheid van hepatocyten voor oxidatieve stress geïnduceerde celdood wordt gepotentialiseerd door lysosomale instabiliteit met verhoogde vrijstelling van ijzer. (14) ROS wordt geacht om ook DNA schade te kunnen induceren. (23)
5) De pathogenese van acute acetaminophen-geinduceerde leverziekte
Bij het onderzoeken van de hepatotoxiciteit van APAP (acetaminophen) werd gezien dat tijdens levermetabolisatie een reactief metaboliet gevormd wordt door het P450 systeem (voornamelijk CYP2E1), namelijk N-acetyl-pbenzoquione imine (NAPQI), wat normaal in de hepatocyt gecapteerd kan worden door GSH. (24) Als bij een overdosis een excessieve
12 hoeveelheid NAPQI ontstaat, gaat dit niet meer gecapteerd kunnen worden en zal het reageren met sulfidryl groepen op eiwitten, leidend tot de vorming van eiwitadducten. Deze adducten kunnen binden op mitochondriale eiwitten en zo het elektronentransport verstoren, waardoor een oxidatieve stress en peroxynitriet in de mitochondriën ontstaan. Hierdoor gaat de membraanpotentiaal verstoord worden wat gaat leiden tot de opening van de MPT-poriën (membrane permeability transition pore), het begin van een necrotische reactie. (25) Necrose leidt vervolgens tot de vrijstelling van DAMPs, die residente immuuncellen (waaronder Kupffer-cellen) zullen activeren om het aangeboren immuunsysteem in het proces te betrekken. (24) Figuur 5: mechanismen van APAP-geïnduceerde necrose [overgenomen uit (25)]
P450 metabolisatie van acetaminophen (APAP) leidt tot de vorming van NAPQI, wat eiwit-adducten met mitochondriale eiwitten tot stand brengt en oxidatieve celstress induceert. Peroxynitriet wordt gevormd uit de reactie van superoxide met NO (stikstofmonoxide). SOD2 (superoxide dismutase 2) kan superoxide omzetten tot waterstof peroxide, maar peroxynitriet kan SOD2 inactiveren. De initiële oxidatieve celstress kan ook het MAP kinase c-jun-N-terminal kinase (JNK) activeren via verschillende redox-gevoelige kinases, waardoor dit JNK gefosforyleerd wordt, leidend tot een translocatie naar de mitochondriën. Deze translocatie van JNK amplificeert de oxidatieve stress, leidend tot MPT porie opening, terwijl apoptotische eiwitten naar de celkern getransloceerd worden. DNA-fragmentatie treedt op en oncotische necrose treedt in werking. Mitofagie is een verdedigingsmechanisme gelijkaardig aan autofagie waarmee beschadigde mitochondriën verwijderd kunnen worden. Afkortingen: ASK-1, apoptosis signal regulating kinase-1; Mkp-1, mitogen-activated protein kinase (MAPK) phosphatase (Mkp)-1; MLK-3, mixed-lineage kinase 3; PTP1B, protein tyrosine phosphatase 1B; RIP
Acetaminophen geïnduceerde hepatotoxiciteit leidt zoals beschreven tot necrotische celschade, wat gepaard gaat met de vrijstelling van DAMPs (damage associated molecular patterns). Bij APAP-hepatotoxiciteit zijn dit voornamelijk HMGB1 (high-mobility group box 1), HSPs (heat shock proteins), DNA fragmenten, mitochondriaal DNA en andere. (24) Residente immuuncellen in de lever, waaronder Kupffer cellen, hebben PRRs voor verschillende DAMPs (TLRs, purinerge receptor P2X7), waarbij na receptor-activatie deze cellen een verhoogde expressie van chemokines en cytokines vertonen, met als doel een pro-inflammatoire reactie op gang te brengen en andere immuuncellen te recruteren. (26) Ook inflammasoom-activatie speelt hier een rol. (27) Tot de gerekruteerde cellen behoren neutrofielen, maar ook eosinofielen, dendritische cellen, NK(T) en T-cellen. (28)
13 De hepatotoxiciteit van APAP is dosis afhankelijk, en in zijn therapeutische dosis is het een veilig middel. (25) Vaak wordt APAP gebruikt in zelfmoordpogingen, maar er wordt een toenemende trend gezien in niet-intentionele overdosissen, wat onder andere verklaard kan worden door de aanwezigheid van het product in preparaten die meerdere geneesmiddelen bevatten. (24) Risicofactoren om sneller te evolueren naar een beeld van ALF hebben te maken met de GSH (glutathione) reserves, waardoor malnutritie, ondervoeding en chronische leverziekten voorbeschikkende factoren zijn. Ook geneesmiddelen die CYP-450 induceren, zoals isoniazide, fenobarbital en rifampicine, alsook chronisch alcoholmisbruik leiden tot een verhoogde productie van NAPQI (N-acetyl-pbenzoquione-imine), het reactieve metaboliet van APAP. (29)
6) Acute leverziekte: huidige aanpak en therapiemogelijkheden
Bij de behandeling van acute leverziekte met als ergste variant ALF is het van belang via uitgebreid onderzoek een oorzaak te achterhalen en zo een oorzaak-specifieke behandeling in te stellen, in combinatie met standaard intensive care om de patiënt te ondersteunen en zo leverregeneratie te optimaliseren, terwijl complicaties voorkomen moeten worden. De intensive care ondersteuning van ALF patiënten vertoont overeenkomsten met de standaard richtlijnen voor sepsis en acute hersenschade. Een oorzaak-specifieke behandeling wordt best zo vroeg mogelijk gestart, en de effecten moeten regelmatig geëvalueerd worden zodat een ELT-indicatie (emergency liver transplantation) bij een risico-patiënt ongeremd gesteld kan worden. (2, 6)
Leverziekte – en falen hebben verschillende gekende complicaties. Cardiovasculaire dysfunctie en hypotensie treden regelmatig op. (3) Zoals kort eerder aangehaald, is ook een verhoogde vatbaarheid voor infecties met risico op sepsis een belangrijke mortaliteitsoorzaak bij acute leverziekte. Een verhoogde permeabilisatie van de intestinale tractus alsook immuundysfunctie staan hierbij aan de basis. SIRS (systemic inflammatory response syndrome) komt regelmatig voor, en dit syndroom is het gevolg van de systemische aanwezigheid van pro- of anti- inflammatoire cytokines. SIRS is geassocieerd met een verhoogde mortaliteit en de ernst van orgaanfalen, en wordt verder geacht een belangrijke factor te zijn in het ontwikkelen van nierfalen en hepatische encefalopathie. Als reactie op SIRS wordt ook vaak een CARS (compensatory anti-inflammatory response syndrome) gezien, wat gekenmerkt wordt door een verhoogde aanwezigheid van anti-inflammatoire mediatoren. (20) Er is reeds gesuggereerd dat het meten van pro-inflammatoire cytokines van belang kan zijn bij het inschatten van de prognose, en overwogen moet worden om deze in selectiecriteria op te nemen. (30)
Encefalopathie is door de progressie tot cerebraal oedeem en intracraniële hypertensie (ICH) gevreesd bij patiënten met ALF. (6) De pathofysiologie is onvolledig begrepen, maar een
14 centrale rol wordt erkend voor ammonium en inflammatoire cytokines. (31) Tegenwoordig probeert men de ammonium-concentratie te verlagen via extracorporele verwijdering via nierfunctievervangende therapie of via therapeutische hypothermie, wat via een vertraging van het lichaamsmetabolisme de productie en cerebrale opname van ammonium zou reduceren. (2)
In het geval van acute APAP geïnduceerde leverziekte, is N-acetylcysteine (NAC) een van de weinige erkende redmiddelen bij een overdosis, waarbij het therapeutisch effect het grootste is als het binnen de acht uur na de overdosis toegediend wordt. Ook na 24u kan een NAC toediening nuttig zijn, maar het effect van de therapie zal veel geringer zijn, en het risico op ALF bijgevolg hoger. (25) Mechanistisch gaat NAC de hepatische synthese van GSH opdrijven door cysteine te voorzien, waardoor meer NAPQI gecapteerd kan worden en minder eiwit-adducten gevormd worden. NAC op zich is ook in staat om toxische metabolieten of radicalen te capteren. Ook leidt het tot verhoogde bloedtoevoer richting de lever en verhoogde mitochondriale ATP productie. (32)
Als laatste redmiddel is er de ELT (emergency liver transplantation), waarvoor een juiste indicatie cruciaal is. Wereldwijd worden hiervoor verschillende selectiecriteria gebruikt, die proberen de prognose in te schatten. Veelgebruikt zijn de King’s College Criteria (KCC). (2) Echter, deze criteria zijn continu in evolutie, naargelang resultaten van overleving na ELT gepubliceerd worden en de criteria regelmatig in vraag gesteld worden. (1) ELT wordt beschouwd als de rescue voor ALF-patiënten die het waarschijnlijk niet zouden overleven met medische zorg alleen. Een analyse van de European Liver Transplantation Registry gaf aan dat 8% van alle levertransplantaties te wijten zijn aan ALF als primaire indicatie. (5)
Voor ELT als maatregel gehanteerd werd, was de mortaliteit bij ALF groter dan 80%. Sinds de invoering van ELT is de overleving enkel maar gestegen, met een overall overleving van 70% en een 2-jaars overleving van 92,4 % voor zij die een levertransplantatie kregen. (4)
15
De neutrofiel in acute leverziekte
1) De neutrofiel in het immuunsysteem
Na de activatie van het immuunsysteem door PAMPs of DAMPs worden verschillende immuuncellen naar de ontstekingshaard geleid, waarvan neutrofielen als eerste gerekruteerd worden. (11)
Neutrofielen (PMN; polymorphonucleaire leukocyten) zijn afkomstig uit het beenmerg en zijn kortlevend (12-18u halfleven) in het perifere bloed, en hun aantal en levensduur is beïnvloedbaar door cytokines als granulocyte colony-stimulating-factor (G-CSF) en granulocyte macrophage colony-stimulating-factor. (33) Hun functie bij inflammatie bestaat uit de snelle eliminatie van micro-organismen en het verwijderen van dode en stervende cellen, opdat regeneratie kan optreden. Echter, een excessieve inflammatoire reactie van neutrofielen kan leiden tot bijkomende schade, wat onder andere in de lever reeds aangetoond is in verschillende experimentele modellen. (34)
Neutrofiel-activatie zou bijgevolg beschouwd kunnen worden als een langs twee kanten snijdend zwaard: cruciaal voor verdediging tegen infecties en regeneratie mogelijk te maken, maar potentieel gevaarlijk door een mogelijke exacerbatie van de oorspronkelijke schade.
2) De neutrofiel in de lever
Neutrofiel-werking in de lever vereist verschillende stappen: rekrutering in de vasculatuur van de lever, extravasatie naar het leverparenchym en een adherentie-dependente cytotoxiciteit. (35)
a) Rekrutering naar de levervasculatuur
De rekrutering naar de levervasculatuur wordt gestart na blootstelling aan inflammatoire mediatoren, zoals TNF-α, IL-1α, IL-1β, CXC chemokines (zoals IL-8, KC (keratinocyte-derived-cytokine; muriene variant van CXCL1), MIP-2 (macrophage inflammatory protein-2; muriene variant van CXCL2), CINC-1 (cytokine-induced neutrophil chemoattractant; variant van IL-8 in ratten)), geactiveerde complementfactoren zoals C5a en andere. Ook DAMPs kunnen
chemotactische effecten op neutrofielen uitoefenen. (34)
Neutrofielen brengen meer dan 30 verschillende receptoren tot expressie die de migratie en het gedrag van de neutrofiel kunnen beïnvloeden, en ze kunnen dus door een rijke variatie aan molecules geactiveerd of gemoduleerd worden, waaronder ook G-CSF en interferonen. (36) Neutrofielen brengen onder andere de meeste TLRs tot expressie (uitgezonderd TLR3 en TLR7, die viraal RNA herkennen), maar ze hebben ook andere PRRs, waarmee ze bijvoorbeeld N-formyl peptiden kunnen herkennen, wat zowel uit mitochondriën als bacteriën
16 kan komen. Dichtbije neutrofielen worden eerder door DAMPs geactiveerd, terwijl chemokines vooral de verdere neutrofielen gaan activeren tot migratie. (37)
Na activatie van de neutrofiel wordt de expressie van adhesiemoleculen op het oppervlak van de neutrofiel verhoogd, zoals CD11b/CD18 (β2-integrine), en de neutrofiel bereidt zich voor op ROS formatie door delen van het NADPH oxidase naar het oppervlak te brengen. Tijdens de rekrutering veroorzaken neutrofielen geen schade; extravasatie in het leverparenchym lijkt een obligate stap te zijn voor cytotoxiciteit uit te oefenen. (34) Waar accumulatie in de (postsinusoidale en portale) venules afhankelijk is van adhesiemolecules (selectines en integrines), verloopt accumulatie in de leversinusoïden via een alternatief mechanisme, namelijk de interactie tussen CD44 en hyaluronan. (38) Accumulatie in de postsinusoidale of portale venules is afhankelijk van de binding van β2-integrine met counterreceptor op het endotheel ICAM-1 (intercellular adhesion molecule-1) of β2-integrine met counterreceptor VCAM-1 (vascular adhesion molecule-1). Belangrijk is de stelling dat hepatotoxiciteit meestal veroorzaakt wordt door neutrofielen die extravaseren vanuit de sinusoïden. (34, 35)
b) Extravasatie naar het leverparenchym
Extravasatie naar het parenchym begint met een sterke adhesie aan het endotheel, waarna de neutrofiel via de exocytose van zijn gelatinase granules matrix metalloproteinases loslaat, die de migratie van de neutrofiel bevorderen door de extracellulaire matrix te degraderen. Echter, bij ernstige endotheelschade kan een adhesie-onafhankelijke transmigratie/extravasatie optreden. (35)
Voor extravasatie is een signaal vanuit het parenchym obligaat. Onder bepaalde omstandigheden zijn CXC chemokines potente stimuli voor extravasatie te bewerkstelligen, maar enkel wanneer zij op het juiste moment op de juiste plaats met een adequate gradiënt vrijgesteld worden. Necrose veroorzaakt ook extravasatie van neutrofielen, waarschijnlijk via complementactivatie na het vrijkomen van verschillende DAMPs en via ROS producten, zoals LPO producten die ook chemotactisch werken voor neutrofielen. (34) Neutrofielen zelf zijn in een tweede fase in staat de initiële rekrutering te versterken door meerdere neutrofielen aan te trekken, via de aanmaak van potente chemotactische factoren zoals leukotrien B4 (LTB4) en
CXCL8. (37)
Figuur 6: fases van neutrofiel rekrutering. [Overgenomen uit (36)]: DAMPs en andere signalen activeren de rekrutering van dichtbije neutrofielen (a) Als reactie op DAMPs, gaan de residente levercellen chemokines en leukotriënen produceren om verdere rekrutering te bewerkstelligen. (b) Gearriveerde neutrofielen gaan direct en indirect de rekrutering onderhouden via de productie van chemokines en leukotriënen, en zo wordt de inflammatie geamplificeerd. (c) In een infectieus milieu zijn er extra signalen die op neutrofielen inwerken en de rekrutering versterken, waaronder PAMPs en signalen gesecreteerd door andere immuuncellen (op de figuur T-cellen, Kupffer cellen en gerekruteerde monocyten/macrofagen). (d)
17
c) Werking van neutrofielen in de lever
Na extravasatie kunnen neutrofielen kleinere molecules fagocyteren of binden aan grotere doelcellen (waaronder hepatocyten) op een selectine (ICAM-1)/integrine dependente manier. Binding aan hepatocyten start een adherentie-dependente oxidatieve stress en degranulatie van de neutrofiel. Hepatocyten zijn geen passief doel voor de neutrofiel: de expressie van ICAM-1 en de aanmaak van chemokines door hepatocyten is onderhevig aan de blootstelling aan inflammatoire cytokines. (35) De oxidatieve stress geïnduceerd door de neutrofiel verloopt als volgt: NADPH oxidase van de neutrofiel genereert superoxide, wat leidt tot de vorming van waterstofperoxide (H2O2), wat gemakkelijk in de hepatocyt kan diffunderen. Ook wordt uit de
azurofiele granules myeloperoxidase vrijgesteld, wat hypochlorigzuur (HClO) kan produceren, wat een vlot diffundeerbare molecule is. (34)
De vier granules van neutrofielen bevatten een repertoire aan molecules die zowel pro-als anti-inflammatoire karakteristieken vertonen. De primaire (azurofiele) granules bevatten factoren zoals MPO en neutrofiel elastase (NE). Secundaire (specifieke) granules bevatten onder andere het PRGP (peptidoglycan recognition protein), M-ficolin en lactoferrine, belangrijke antibacteriële producten. De tertiaire (gelatinase) granules bevatten matrix degraderende molecules, zoals MMP (matrix metalloprotease), en deze worden vrijgelaten bij transmigratie van de neutrofiel, volgend op de binding van integrines op selectines op het endotheel. De secretorische vesikels bevatten onder andere membraan geassocieerde molecules als complement receptoren, LPS binding receptor, componenten van het NADPH oxidase, β2 integrines (CD11b/CD18) en andere, en worden losgelaten bij de vrijstelling aan inflammatoire mediatoren, met als doel de neutrofiel klaar te maken voor verdere migratie en
18 adherentie. Bij het binden van de neutrofiel aan zijn doel (hepatocyt, micro-organisme), worden de azurofiele en specifieke granules losgelaten, met vrijstelling van bactericiede producten (defensines etc) en serine proteases, waaronder neutrofiel elastase, cathepsine G en proteinase-3, alsook MPO wat belangrijk is voor het genereren van ROS. (34, 37)
Serine proteases (waaronder neutrofiel elastase; NE) afkomstig uit neutrofielen zijn in staat om directe hepatotoxiciteit te veroorzaken, en ROS wordt geacht mede verantwoordelijk te zijn voor deze vorm van celdood, omdat een oxidatieve stress serine-protease-inhibitoren zoals α1-antitrypsine, α2 macroglubuline en α1-antichymotrypsine inhibeert, terwijl de serine proteases zelf minder onderhevig zijn aan deze oxidatieve stress. Hierdoor ontstaat een regio rond de neutrofiel waarbinnen serine proteases actief kunnen zijn. Echter, serine proteases hebben nog meerdere functies inzake de regulatie van het inflammatoire proces, waaronder de vrijstelling van TNF-α en de activatie van TGF-α (tissue growth factor). Elastase kan ook Kupffer cellen induceren tot de vorming van chemo-attractieve molecules voor monocyten en neutrofielen. Grofweg kunnen de functies van serine proteases naast potentiele hepatotoxiciteit onderverdeeld worden in de activatie van receptoren, het verwerken en loslaten van pro-inflammatoire cytokines en modulatie van de activiteit van cytokines en chemokines. (34, 39)
Neutrofielen zijn niet enkel effectorcellen van het immuunsysteem, maar hebben een bredere rol in inflammatie en regeneratie, met beïnvloeding en rekrutering van andere immuuncellen door productie van chemokines en cytokines, alsook via directe interactie met macrofagen, dendritische cellen, lymfocyten en NK cellen. (40) Bijvoorbeeld, de differentiatie van gerekruteerde monocyten tot macrofagen wordt gestuurd door chemokines, pro- en anti-inflammatoire cytokines (zoals IL-8 en IL-10) en vetmolecules gesecreteerd door neutrofielen. IFN-γ uit neutrofielen helpt met het rekruteren van macrofagen, en G-CSF uit neutrofielen versterkt de aanwezigheid en aanmaak van neutrofielen. Het is aangetoond dat neutrofielen zelf gepolariseerd (N1 als pro-inflammatoir fenotype en N2 als anti-inflammatoir fenotype) kunnen worden als respons op bepaalde stimuli, wat grote effecten op het verloop van de inflammatie teweeg brengt, en de immunomodulerende functie van neutrofielen bevestigt. (37) N1 neutrofielen (pro-inflammatoir) brengen vele pro-inflammatoire cytokines en chemokines tot expressie, terwijl N2 neutrofielen (anti-inflammatoir) eerder gekenmerkt worden door hoge expressie van CXCR4 (CXC-chemokine receptor 4), VEGF (vascular endothelial growth factor) en MMP-9 (matrix metalloproteinase 9). TGF-β (transforming growth factor) lijkt bepalend te zijn voor welk fenotype zich preferentieel ontwikkelt. (36)
19 Figuur 7: interactie van de neutrofiel met de omgeving en het immuunsysteem. [Overgenomen uit (37)]
NETosis (neutrophil extracellular traps) is een gespecialiseerde vorm van celdood, met expulsie van chromatinedraden en histonen die gecoat zijn met allerlei producten uit de granules van neutrofielen, en wordt geactiveerd door contact met bepaalde cytokines, micro-organismen of producten van micro-micro-organismen. (37) Ook vrije vetzuren kunnen NETosis uitlokken. (41) De webachtige structuur die uitgestoten wordt, kan pathogenen immobiliseren met een versterkte anti-microbiële werking door het pathogeen te verplichten in contact te komen met ROS, lytische enzymes en anti-microbiële agentia, met verminderde schade aan de omgeving. (37)
d) De neutrofiel en resolutie van inflammatie
Om verdere celschade te vermijden, is het nodig dat neutrofielen tijdig verwijderd worden uit de inflammatiezone. Dit kan gebeuren via apoptose of necrose of fagocytose door macrofagen. (36) Macrofagen worden door neutrofielen naar de inflammatoire zone geleid via de loslating van MIP-1α (macrophage inflammatory protein-1α), MIP-1β en IFN-γ, waar ze apoptotische en non-apoptotische neutrofielen kunnen fagocyteren, alsook MPO. Afhankelijk van wat de macrofaag opneemt en de status van de inflammatie, gaat deze een M1 (pro-inflammatoir) of M2 (pro-resolutie) fenotype aannemen, wat cruciaal is voor het onderhouden of termineren van de inflammatie. Het M2 fenotype is geassocieerd met een productie van anti-inflammatoire mediatoren (zoals IL-10), het fagocyteren van apoptotische neutrofielen en
20 een loslating van defensines, resolvines en MMP-9, belangrijk voor neo-angiogenese. Het verwijderen van neutrofielen is belangrijk om resolutie toe te laten, opdat de macrofaag de dominantere cel wordt en er ruimte is voor de rekrutering van onder andere fibroblasten voor de heropbouw van de matrix. (37)
In verschillende modellen (zebravis, muizen en in vitro menselijke neutrofielen) is aangetoond dat neutrofielen niet hoeven te sterven, maar de inflammatiezone kunnen verlaten via een proces genoemd ‘reverse migration’ en ‘neutrophil reverse transendothelial migration’, respectievelijk interstitiële migratie van neutrofielen weg van de ontstekingszone en terug toetreden van neutrofielen tot de vasculatuur. (36)
Vraagstelling
De therapeutische opties bij acute leverziekte zijn eerder beperkt, waardoor bij evolutie tot ALF soms overgegaan moet worden tot een ELT om het leven van de patiënt te redden. Om de transplantatienood te beperken, is het nodig dat gezocht wordt naar nieuwe strategieën om leverschade te beperken. Inflammatie staat centraal in de pathogenese van acute leverziekte, en dit kan dus een interessant therapeutisch doel zijn om de schade te beperken. In deze masterproef zal de rol van neutrofielen in een model van paracetamol-geinduceerde hepatotoxiciteit onderzocht worden, wat epidemiologisch gezien in de Westerse wereld de frequentste etiologie is. Gerapporteerde interventies tegen deze groep cellen zullen onderzocht worden om te kijken of op dit niveau bij paracetamol geinduceerd leverfalen therapeutische winst geboekt kan worden ter vermijding van de progressie tot ALF.
21
Methodologie
Voor het uitvoeren van deze literatuurstudie, werd gewerkt in verschillende fases. Enkel artikels in het Engels werden geïncludeerd.
De eerste fase bestond uit de oriëntatie in het onderzoeksveld. Voor deze fase werden artikels gezocht vanaf 2005, waarbij de laatste zoekopdracht verricht werd op 01/10/2019. Gebruik makend van MEDLINE en Google Scholar werd gezocht naar de definitie, etiologie, epidemiologie en aanpak van acute leverziekte en acuut leverfalen (ALF). Er werd gezocht met de termen (MeSH en vrije tekst) ‘acute liver failure’, ‘acute liver disease’ en ‘liver injury’. Om de waarde van de gevonden artikels in te schatten, werd gekeken naar de impact factor van de gevonden artikels. Vervolgens werden aangehaalde termen gecombineerd met specifiekere termen en subheadings, waaronder ‘epidemiology’, ‘etiology’, ‘therapy` en ‘management`. Om inzicht te verkrijgen in de pathogenese van acute leverziekte, werd nogmaals gezocht met de termen ‘acute liver failure’, ‘acute liver disease’ en ‘liver injury’, nu gecombineerd met de zoektermen ‘inflammation’, ‘pathogenesis’, ‘infection’ en ‘immunity’. Hierna werd dit proces herhaald, nu met toegevoegde specifiekere zoektermen om in de inleiding aangehaalde mechanismen beter te begrijpen: ‘apoptosis’, ‘necrosis’, ‘cell death’, ‘toll like receptors’, ‘reactive oxygen species’, ‘pyroptosis` en ‘inflammasome’.
Hierna werd de focus op de neutrofiel verlegd, met als eerste doel de werking van de neutrofiel in inflammatie en vervolgens in de lever te verduidelijken. Eerst werd gezocht met termen ‘neutrophil function’, `neutrophil infiltration`, ‘neutrophil induced injury’, ‘neutrophil hepatotoxicity’ en ‘neutrophil inflammation’. Omdat snel duidelijk werd dat dit een te brede selectie aan artikels opleverde, werd gecombineerd met de termen ‘acute liver failure’, ‘acute liver disease’ en ‘liver injury’, wat relevantere artikels opleverde.
Bij de zoektocht naar artikels om de onderzoeksvraag te beantwoorden, namelijk de rol van neutrofielen in APAP-geinduceerde acute leverziekte, werd gebruik gemaakt van de databank van MEDLINE. Er werd gezocht vanaf 2000, waarbij de laatste zoekopdracht werd verricht op 14/11/2019. Artikels die niet in het Engels geschreven zijn werden geexcludeerd. Er werd gezocht met de termen `neutrophils` of `polymorphonucleair leukocyt` (als MesH term en als vrije tekst), gecombineerd met vrije zoektermen `acetaminophen` of `APAP`, alsook met de termen `liver failure`, `hepatotoxicity` of `liver injury`. Gebaseerd op de abstracts werden artikels geïncludeerd wanneer er onderzocht werd in een model van acetaminophen hepatotoxiciteit, en wanneer de functie van neutrofielen of de cytokines/chemokines betrokken bij neutrofielfunctie onderzocht werden. Enkel artikels waarvoor de volledige tekst online beschikbaar was werden geïncludeerd.
22
Resultaten
1) Argumenten voor een rol van neutrofielen in het exacerberen van APAP
geïnduceerde leverschade
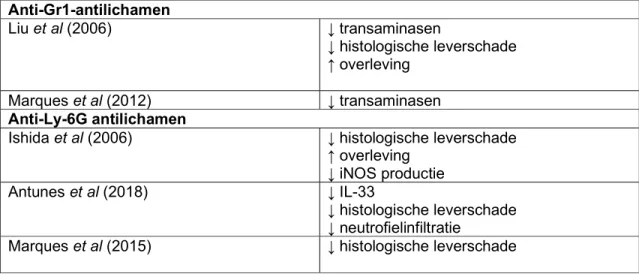
Liu et al toonden in 2006 aan dat een anti-Gr-1 (anti-neutrofiel) antilichaam gegeven aan muizen 24u en 4 uur voor ze een APAP-overdosis (500mg/kg) kregen leidde tot leverbescherming, gekwantificeerd via het ALT en AST-gehalte in het bloed (sALT), necrotische zones op leverbiopt en de overleving van de muizen ten opzichte van de controles. Ook rapporteerden zij dat anti-Gr-1 antilichaam neutrofiel-accumulatie in de lever kon tegengaan, zonder dat dit significant de rekrutering van andere leukocyten en immuuncellen verminderde (NK cellen, NKT cellen, T cellen en macrofagen). Om de rol van ICAM-1 in APAP-hepatotoxiciteit te evalueren, werd gekeken naar leverschade en neutrofielinfiltratie bij ICAM-1 deficiënte muizen. Bij ICAM-ICAM-1 deficiënte muizen werd een verminderde rekrutering van neutrofielen naar de lever gevonden na APAP-intoxicatie, en dit ging gepaard met een verminderde histologische leverschade wanneer vergeleken met het wild type. (42)
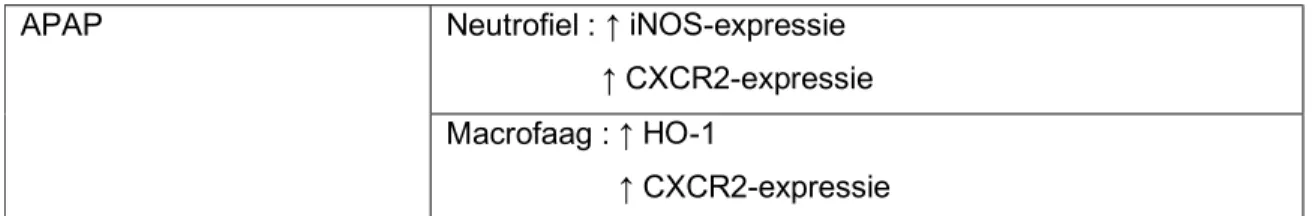
Ishida et al onderzochten APAP-geïnduceerde leverschade door 750mg/kg APAP intraperitoneaal toe te dienen bij muizen. Een eerste bevinding was dat APAP-toediening leidde tot een verhoogde expressie van iNOS (inducible NO synthase; draagt bij tot verhoging van oxidatieve stress; zie supra) en HO-1 (haem oxygenase) in het leverparenchym, en dat dit tot uiting gebracht werd door respectievelijk neutrofielen en macrofagen in de lever. HO-1 zou protectief zijn in de pathogenese; HO-1 inhibitie leidde tot een significante verhoging van de mortaliteit. Ook ging APAP-intoxicatie gepaard met een verhoogde hepatische expressie van CXCR2, zowel door neutrofielen als macrofagen. Wanneer de muizen behandeld werden met een anti Ly-6G granulocyt antilichaam (veroorzaakt neutropenie), leidde dit tot een verminderde leverschade, een verbeterde overleving en verminderde neutrofiel-rekrutering na APAP-toediening, alsook een verminderde iNOS productie, wat samen lijkt te suggereren dat neutrofielen een pathologische rol spelen in het ziekteproces. (43)
Tabel 2 : hypothese van Ishida et al (43) : zowel neutrofielen als macrofagen worden geactiveerd tot rekrutering richting het necrotische APAP-leverparenchym, waarbij de outcome van het proces mede bepaald wordt door het evenwicht tussen beschermende (HO-1) macrofagen en neutrofielen die via iNOS de schade kunnen verergeren
APAP Neutrofiel : ↑ iNOS-expressie
↑ CXCR2-expressie Macrofaag : ↑ HO-1
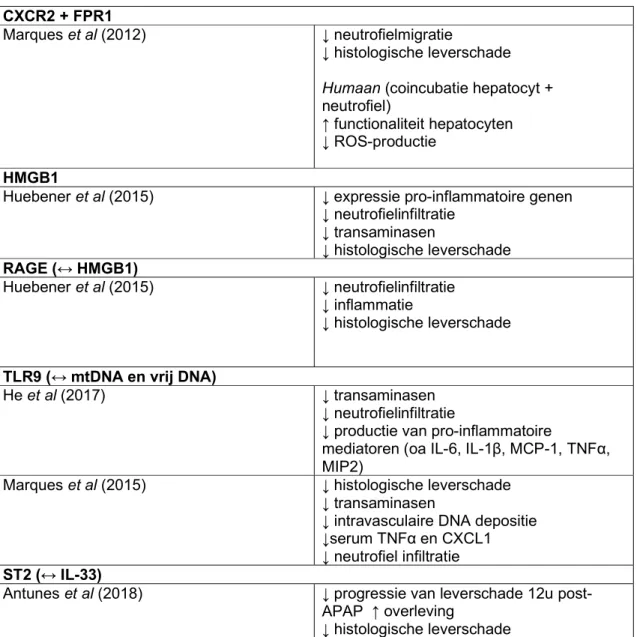
23 Marques et al onderzochten in 2012 de rol van CXCR2 chemokines en mitochondriale producten tijdens APAP-hepatotoxiciteit, alsook het effect op neutrofiel-rekrutering naar de lever en geassocieerde systemische inflammatie en leverschade. Om te bepalen of neutrofielen bijdroegen tot de schade, gebruikten de onderzoekers verschillende methoden: allereerst werd een anti-Gr-1 antilichaam toegediend met als doel neutrofiel-depletie te bekomen, en er werd gezien dat dit leidde tot een reductie in sALT waarden wanneer vergeleken met controles (bij muizen). Ten tweede werden HepG2 cellen (humane hepatocyten) samen met humane neutrofielen geïncubeerd in verschillende proporties (1:1 en 10:1 HepG2 cellen/neutrofielen), en er werd gezien dat de toevoeging van neutrofielen de levensvatbaarheid (op basis van het metabolisme) van de HepG2 cellen verstoorde, en wanneer beide celtypes gescheiden werden in een transwell-coculture systeem verminderde de eerder waargenomen cytotoxiciteit significant. Wanneer humane neutrofielen toegevoegd werden aan APAP-geintoxiceerde HepG2-cellen (in analoge verhoudingen als bij de eerste coincubatie), was de geobserveerde celdood en ROS-productie significant verhoogd wanneer vergeleken met niet-APAP voorbehandelde HepG2 cellen. Verder toonde een ELISA-analyse van muriene post-APAP levers een verhoogde expressie van CXCR2 chemokines (CXCL1 en CXCL2) 24u na APAP-toediening, en een analoog resultaat werd gezien voor CXCL8 bij de HepG2 cellen na APAP-toediening. CXCR2 blokkade in het muismodel leidde tot een gedeeltelijke reductie in neutrofiel-infiltratie, maar geen bescherming tegen leverschade werd vastgesteld. DNA fragmenten en mitochondriale producten zoals formyl peptiden kunnen neutrofielen aantrekken en activeren via respectievelijke receptoren TLR9 en FPR1. Wanneer CXCR2 samen met FPR1 geblokkeerd werden, leidde dit tot een significante inhibitie van neutrofiel-migratie en leverschade (bij muizen), en in het HepG2-neutrofiel coincubatie model leidde deze blokkade tot een significante inhibitie van cytotoxiciteit en ROS-productie, wat een bijdrage van de FPR1-CXCR2 pathway in neutrofiel-geïnduceerde hepatotoxiciteit lijkt te bevestigen. (44)
Huebener et al evalueerden de rol van HMGB1 in de context van necrose, in een transgeen muismodel waarbij het HMGB1-coderend gen geableerd werd. Muizen werden genetisch gecodeerd met langs de ene kant volledige HMGB1 deficiëntie (HMGB1KO), en langs de andere kant met selectieve hepatische HMGB1 deficiëntie (HMGB1hepKO). Na APAP-toediening was de leverschade vergelijkbaar tussen de HMGB1KO en HMGB1hepKO groepen, maar een sterke reductie van vroege en late neutrofielinfiltratie en expressie van pro-inflammatoire genen werd gezien bij de HMGB1hepKO groep. HMGB1 deletie had geen effecten op de infiltratie van macrofagen. Ook werd gezien dat naast een verminderde neutrofielinfiltratie bij HMGB1hepKO muizen een verminderde leverschade aanwezig was, zoals geëvalueerd door sALT/sAST en histologische levernecrose. Als receptor voor HMGB1
24 op neutrofielen identificeerden de onderzoekers RAGE (receptor for advanced glycation end-products), omdat de migratie van neutrofielen verstoord was bij RAGE-deficiënte muizen, en zij zagen ook dat neutrofiel-infiltratie, inflammatie en leverschade verminderd waren bij RAGE-deficiënte muizen. Vervolgens onderzochten ze neutrofiel-geïnduceerde schade bij APAP-cytotoxiciteit, door muizen te coderen die in het beenmerg deficiënt waren in het neutrofiel-elastase (NE). Deze NE-deficiënte muizen vertoonden een verminderde necrose en sALT-niveaus na APAP-toediening. Als aanvulling werd de impact van een HMGB1-inhibitor (glycyrrhizin) geëvalueerd, en dit zou APAP-geïnduceerde hepatotoxiciteit verminderen in een gelijkaardige kwantiteit als genetische HMGB1 ablatie, al was dit effect het meest prominent als de inhibitor voor de APAP-toediening toegediend werd. (45)
Tabel 3: gerapporteerde effecten van interventies tegen chemokines, cytokines en DAMPs of hun receptoren betrokken bij neutrofielfunctie tijdens APAP-hepatotoxiciteit pleitend voor een
exacerberende rol van neutrofielen in de pathogenese CXCR2 + FPR1
Marques et al (2012) ↓ neutrofielmigratie
↓ histologische leverschade Humaan (coincubatie hepatocyt + neutrofiel)
↑ functionaliteit hepatocyten ↓ ROS-productie
HMGB1
Huebener et al (2015) ↓ expressie pro-inflammatoire genen ↓ neutrofielinfiltratie ↓ transaminasen ↓ histologische leverschade RAGE (↔ HMGB1) Huebener et al (2015) ↓ neutrofielinfiltratie ↓ inflammatie ↓ histologische leverschade
TLR9 (↔ mtDNA en vrij DNA)
He et al (2017) ↓ transaminasen
↓ neutrofielinfiltratie
↓ productie van pro-inflammatoire
mediatoren (oa IL-6, IL-1β, MCP-1, TNFα, MIP2)
Marques et al (2015) ↓ histologische leverschade ↓ transaminasen
↓ intravasculaire DNA depositie ↓serum TNFα en CXCL1 ↓ neutrofiel infiltratie ST2 (↔ IL-33)
Antunes et al (2018) ↓ progressie van leverschade 12u post-APAP ↑ overleving
25 ↑ recuperatie ↓ neutrofielinfiltratie ↓ CXCL1/2 in serum CXCL16 Wang et al (2017) ↓ transaminasen ↓ HMGB1 in serum ↓ histologische leverschade ↑ overleving ↓ oxidatieve stress ↓ neutrofielinfiltratie ↓ CXCL1 en CXCL2 mRNA expressie ↓ ICAM-1 expressie
↓ JNK fosforylatie (betrokken bij ROS-celdood)
↓ hepatische TNFα en IL6 aanwezigheid IL-17
Lee et al (2018) ↓ transaminasen
↓ neutrofielinfiltratie en activiteit (MPO-activiteit)
↓ serum IL-6, TNFα, IFN-γ
↓ ERK fosforylatie (betrokken bij ROS-celdood)
P2Y2R (↔ ATP)
Ayata et al (2012) ↑ NFκβ activatie
↑ overleving hepatocyt ↓ serum CXCL1/2
Czepielewski et al onderzochten de effecten van een GRPR-selectieve (gastrin-releasing peptide) antagonist, RC-3095 (RC), op neutrofiel migratie en leverschade, nadat eerder aangetoond was dat GRP chemotactisch werkt voor neutrofielen, in een muismodel na APAP-intoxicatie. Interessant was dat RC-toediening ook de migratie van neutrofielen gestimuleerd door CXCL8 (IL-8) kon verstoren, en een hypothese van de auteurs was dan ook dat de effecten van RC berustte op modulatie van de CXCR2 (receptor voor CXCL1, 2, 5 en 8). RC had geen effect op de migratie van menselijke neutrofielen richting fMLP en LTB4, en er werd ook gezien dat RC geen effect had op de expressie van CXCR2 (terwijl binding van CXCR2 met bv CXCL8 leidde tot internalisatie van de receptor). Vervolgens werd de impact van RC op de leverfunctie in een APAP-muismodel getest, en er werd gezien dat vroege behandeling met RC significant sALT/sAST verlaagde, histologische schade verminderde en dat de leverfunctie hierbij vlugger herstelde (gebaseerd op toediening van ICG/indocyanine green; vlug geklaard uit bloed door de lever) (wanneer vergeleken met controles). 24 uur na APAP toediening werden de muizen microscopisch intravitaal geëvalueerd, en bij RC-behandelde muizen was er in vergelijking met controles minder necrose, waren er minder geïnfiltreerde neutrofielen, waren de aanwezige neutrofielen meer verdeeld doorheen de lever (in tegenstelling tot geconcentreerde lokalisatie in de necrotische zones bij de controles), hadden
26 de neutrofielen een verminderde motiliteit en een verminderde directionaliteit. Verder toonden de onderzoekers aan dat de inhibitie van RC op CXCL8-gemediëerde chemotaxis berust op een MAPK-pathway fosforylatie met effecten op de expressie van adhesie-molecules CD11b en CD66b en bijgevolg migratie. Bij geïsoleerde muis-neutrofielen werd gezien dat RC voorbehandeling de CXCL2 gemedieerde activatie van neutrofielen via opregulatie van CD11b en het verwijderen van L-selectine (CD62L) inhibeert. (46)
Tabel 4: gerapporteerde effecten van interventies tegen adhesiemolecules betrokken bij neutrofielmigratie naar het leverparenchym tijdens APAP-hepatotoxiciteit pleitend voor een exacerberende rol van neutrofielen in de pathogenese
ICAM-1
Liu et al (2006) ↓ histologische leverschade
He et al (2017) ↓ neutrofielinfiltratie
↓ transaminasen
CXCL2/CXCL8-CXCR2 gemedieerde opregulatie van CD11b en CD66b Czepielewski et al (2017) ↓ transaminasen
↑ leverfunctionaliteit
↓ histologische leverschade
↓ neutrofielinfiltratie in necrotische zones E-selectine
Oancea et al (2016) ↓ neutrofielinfiltratie ↓ transaminasen
↓ histologische leverschade ↓ hepatocyt-apoptose
He et al onderzochten het effect van microRNA-223 (miR-223) op neutrofiel activatie en leverschade, nadat mitochondriaal DNA (mtDNA) TLR9 stimuleert tot de activatie van neutrofielen na APAP-intoxicatie. De studie toonde aan dat miR-223 expressie verhoogd was in neutrofielen en hepatocyten na APAP- intoxicatie, en dat miR-223 ook verhoogd was in het serum. Deletie van miR-223 (miR-223 deficiënte muizen tegenover het WT) leidde tot een exacerbatie van APAP-geïnduceerde neutrofiel infiltratie, leverschade en inflammatie, gebaseerd op sALT/sAST, histologische necrose en een verhoogde neutrofiel/macrofaag infiltratie. Ook was de transcriptie van verschillende cytokines en chemokines opgereguleerd bij miR-223KO muizen ten opzichte van het WT na APAP-toediening, namelijk IL-6, IL-1β, TNFα, MCP-1 (monocyt chemotactic protein) en MIP2 (macrophage inflammatory protein). Om verder te bevestigen of neutrofielinfiltratie bijdroeg tot de verhoogde schade bij miR-223KO muizen, werd een bijkomende deletie van het ICAM gen verricht, en er werd gezien dat deze extra deletie zorgde voor verminderde neutrofiel-accumulatie in de lever na APAP in vergelijking met enkel miR-223KO muizen, alsook een verlaagd sALT/sAST in vergelijking met het WT. Verder werden oxidatieve stress-merkers van miR-223KO muizen vergeleken met het WT, en er werd gezien dat er bij de KO groep na APAP toediening een snellere downregulatie van GSH was, dat er meer hepatisch 4-HNE (4-hydroxynonenal; oxidatieve stress marker)
27 aanwezig was, N-nitrotyrosine (RNS gevormd uit superoxide en NO) verhoogd was, iNOS opgereguleerd was, en SOD1 expressie verlaagd was. miR-223 deficiëntie verhoogde dus duidelijk oxidatieve stress na APAP-toediening. Toediening van een TLR9-antagonist attenueerde schade bij miR-223KO en de WT muizen met verminderde sALT, verminderde neutrofielinfiltratie (gekwantificeerd via Ly6G mRNA en MPO-kleuring) en een verminderde productie van pro-inflammatoire mediatoren, wanneer vergeleken met de controles die geen TLR9 antagonist kregen. Wanneer miR-223KO muizen een TLR9 agonist toegediend kregen, leidde dit in neutrofielen tot een verhoogde expressie van inflammatoire cytokines, in vergelijking met het WT. Ook werd de activatie van het NFκβ vergeleken tussen miR-223KO muizen en het WT door de opregulatie en expressie van IKKα (IκB kinase α; betrokken bij de activatiepathway van NFκβ) te vergelijken, en de aanwezigheid van het eiwit in de lever en in neutrofielen was verhoogd bij de miR-223 deficiënte muizen. Verder werden de effecten van een 223 inhibitor en opregulator geëvalueerd bij neutrofielen, en er werd gezien dat miR-223 opregulatie de TLR9-agonist-gemediëerde opregulatie van IL-6, MIP-1α en MIP-1β kon diminueren, én dat miR-223 opregulatie de aanwezigheid van IKKα kon teniet doen bij TLR9-agonist-gemediëerde NFκβ activatie, en als laatste ook de TLR9-TLR9-agonist-gemediëerde activatie van NFκβ p65 kon doen verminderen. De effecten van een miR-223 inhibitor waren exact de omgekeerde van degene die net beschreven zijn. Als laatste werd een relatie aangetoond tussen mtDNA geïnduceerde TLR9 activatie en de opregulatie van miR-223 in neutrofielen, maar dit zou niet het enige mechanisme zijn dat miR-223 kan doen opreguleren, want toevoeging van een TLR9 antagonist kon de miR-223 opregulatie slechts gedeeltelijk verhinderen, en een rol voor IL-6 in de opregulatie van miR-223 werd bijgevolg aangehaald. De auteurs concluderen dat miR-223 regulatie gedeeltelijk afhankelijk was van de mtDNA-gemediëerde activatie van TLR9, en werkt als een negatieve feedback mechanisme voor overdreven neutrofiel-activatie bij APAP-hepatitis te beperken. In conclusie suggereren de auteurs een potentiele exacerberende rol voor neutrofielen in de pathogenese van APAP-hepatotoxiciteit, terwijl ze tegelijkertijd een fysiologisch mechanisme geven waarbij deze exacerbatie geattenueerd wordt. (47)
28 Figuur 8 [overgenomen uit (47)]: Necrotisch vrijgesteld mtDNA activeert NFκβ op een TLR9 afhankelijke manier, leidend tot inflammatie via de opregulatie van (pro-)inflammatoire mediatoren. Tegelijkertijd wordt ook miR-223 expressie verhoogd, met als effect een negatieve feedback op de mtDNA/TLR9 activatie door de activiteit van IKKα te beïnvloeden. Inhibitie van miR-223 leidde tot een verhoogde oxidatieve stress alsook een toegenomen leverschade, gekwantificeerd via de histologie en de verhoging van transaminasen.
Antunes et al onderzochten de rol van de IL-33/ST2 as in de pathogenese van APAP-geïnduceerde leverschade. IL-33 is een cytokine van de IL-1 familie en is een ligand voor de ST2 receptor (suppression of tumorigenicity 2; behoort tot de IL-1 receptor familie; heeft onder andere een membraangebonden en een oplosbare variant), en IL-33 heeft in de celkern de functie om transcriptie te reguleren, met gekende effecten op NFκβ en MAPK activatie. APAP toediening bij ST2 deficiënte (ST2KO) muizen leidde tot een abrupte stop van de progressie van leverschade op 12u na toediening (gebaseerd op sALT/sAST stijgingen), terwijl bij het WT de necrose door bleef gaan tot 24u na APAP toediening, gebaseerd op aanhoudende sALT/AST stijgingen. Ook was de overleving van ST2KO muizen verlengd bij APAP tegenover WT, en ook de histologische vastgestelde necrose was beperkter, suggererend dat ST2KO muizen, desondanks gelijkaardige initiele leverschade als WT, een snellere recuperatie en een mildere leverschade hadden na APAP-intoxicatie. Amplificatie van oorspronkelijke leverschade lijkt dus IL-33 gemedieerd te zijn. Confocale intravitale microscopie toonde vervolgens dat in het WT APAP-toediening leidde tot een massieve rekrutering van neutrofielen in de lever en dat dit vooral gebeurde in vrij DNA-rijke regio`s, terwijl bij ST2KO muizen minder neutrofielen accumuleerden en minder vrij-DNA rijke zones vastgesteld werden, alsook een verminderde aanwezigheid van CXCL1 en CXCL2 in het serum, wat in totaal een rol voor IL-33/ST2 in neutrofiel rekrutering en schade-amplificatie lijkt te bevestigen. ST2 expressie na APAP-toediening was hoger bij niet-parenchymale cellen dan bij hepatocyten, terwijl als bron van IL-33 voornamelijk hepatocyten en niet-parenchymale cellen gevonden werden. Omdat APAP-intoxicatie leidde tot een significante verhoging van de aanwezigheid van IL-33 producerende neutrofielen, werd een relatie tussen neutrofielen met de IL-33/ST2 as gesuggereerd. Als laatste werd aangetoond dat neutrofiel-depletie door middel van een anti-Ly6G antilichaam leidde tot een significante reductie van hepatische en circulerende IL-33 niveau`s, een verminderde leverschade en (logischerwijs)
29 neutrofielinfiltratie (gekwantificeerd via de aanwezigheid van MPO). De auteurs besluiten dat de IL-33/ST2 neutrofiel activatie een belangrijke inflammatoire pathway is in een context van steriele lever inflammatie. (48)
Wang et al onderzochten in 2017 de rol van CXCL16 in de pathogenese van APAP-geïnduceerde leverinflammatie, door de effecten hiervan in een muismodel te evalueren, door CXCL16-deficiënte muizen (CXCL16KO) te vergelijken met het WT. Allereerst werd vastgesteld dat de mRNA expressie van CXCL16 significant opgereguleerd was in long, hart en lever 24u na APAP toediening, en dit correleerde met de serumniveau`s, in vergelijking met onbehandelde controles. Verder waren CXCL16KO muizen beter beschermd tegen APAP-intoxicatie dan het WT, met een verminderde inductie van sALT/sAST, een verbeterde overleving wanneer een lethale dosis (750 mg/kg) toegediend werd, een verminderde histologische necrose en een verminderde loslating van HMGB1 in het serum. Ook de oxidatieve stress in de lever was verminderd bij CXCL16KO muizen in vergelijking met het WT na APAP toediening, wat zich vertaalde in verminderde ROS productie, verminderde MDA-concentratie (product van vetperoxidatie) en een verminderde activiteit van de mitochondrial respiratory chain (MRC). Ook was de neutrofielinfiltratie in het beschadigd leverparenchym significant verminderd in vergelijking met het WT na APAP, met bij de CXCL16KO muizen ook een minder sterke verhoging van CXCL1 en CXCL2 mRNA als reactie op APAP, en geen verhoogde expressie van ICAM-1 (wat wel gezien werd bij het WT na APAP). Als laatste vaststellingen zagen de auteurs dat JNK-fosforylatie (stress geactiveerd kinase dat een rol speelt bij ROS-geïnduceerde celdood) verminderd was bij CXCL16KO muizen in vergelijking met het WT na APAP, alsook dat hepatische TNFα en IL-6 niveau`s verlaagd waren bij de CXCL16KO muizen (in vergelijking met WT; na APAP). (49)
Lee et al onderzochten de rol van IL-17 in APAP-hepatotoxiciteit door IL-17 deficiënte muizen (IL17KO) muizen te vergelijken met het WT, omdat eerder onderzoek een rol voor IL-17 in de rekrutering van immuuncellen aangaf. IL17KO muizen hadden op 16u na APAP toediening een verminderde sALT/sAST alsook histologische necrose in vergelijking met het WT. De MPO activiteit (neutrofiel-activiteit), neutrofiel-accumulatie en γδ T-cel infiltratie was significant verlaagd bij de IL17KO muizen in vergelijking met het WT na APAP-toediening, terwijl macrofaag infiltratie verhoogd was bij het WT. Ook was de aanwezigheid in het serum van IL-6, TNF-α en IFN-γ lager bij IL-17KO muizen dan bij het WT na APAP-toediening. Verder werd gezien dat IL-17 een belangrijke invloed had op ERK-fosforylatie (maakt deel uit van de ROS-geïnduceerde JNK activatie, wat leidt tot mitochondriale dysfunctie en necrose), want deze fosforylatie was sterk verminderd bij IL-17KO muizen. (50)
![Figuur 3: vereenvoudigd schema van celdood in acuut leverfalen. [overgenomen uit (18)]](https://thumb-eu.123doks.com/thumbv2/5doknet/3285386.21796/11.918.143.531.251.506/figuur-vereenvoudigd-schema-celdood-acuut-leverfalen-overgenomen.webp)