Current focus
Alternative food-preservation technologies: efficacy and mechanisms
Beatrice H. Lado, Ahmed E. Yousef *
Department of Food Science and Technology, The Ohio State University, 2015 Fyffe Road, Parker Hall, Columbus, OH 43210, USA
Abstract
High-pressure processing, ionizing radiation, pulsed electric field and ultraviolet radiation are emerging preservation technologies designed to produce safe food, while maintaining its nutritional and sensory qualities. A sigmoid inactivation pattern is observed in most kinetic studies. Damage to cell membranes, enzymes or DNA is the most commonly cited cause of death of microorganisms by alternative preservation technologies. © 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: High pressure; Injury; Irradiation; Kinetics; Non-thermal; Pulsed electric field; Resistance; Ultraviolet
1. Introduction
Consumers are increasingly aware of the health benefits and risks associated with consumption of food. To meet consumers’ expectations, the food industry is devoting considerable resources and expertise to the production of wholesome and safe products. Production of safe food includes scrutinizing materials entering the food chain, suppressing microbial growth (e.g. storage at chilling tem-perature), and reducing or eliminating the microbial load by processing and preventing post-processing contamination. The presence of a processing unit operation aiming at microbial destruction is of primary importance to ascertain safety and stability of food. Heat treatments are traditionally applied to pasteurize or sterilize food, generally at the expense of its sensory and nutritional qualities. As consum-ers increasingly perceive fresh food as healthier than heat-treated food, the industry is now seeking alternative tech-nologies to maintain most of the fresh attributes, safety and storage stability of food [1].
Satisfactory evaluation of a new preservation technology depends on reliable estimation of its efficacy against patho-genic and spoilage food-borne microorganisms. Research on alternative technologies was initially focused on process design, product characteristics and kinetics of microbial inactivation. The success of these new technologies, how-ever, depends on progress in understanding microbial physi-ology and behavior of microbial cells during and after
treatment. Consequently, this article reviews alternative preservation technologies with emphasis on (i) comparing their efficacy with conventional heat treatment, (ii) mecha-nisms of microbial inactivation, (iii) patterns of inactivation kinetics, (iv) microbial resistance mechanisms and (v) potential causes of underestimation of survivors during food processing by alternative technologies.
2. Food-preservation technologies
Thermal pasteurization and sterilization are predomi-nantly used in the food industry for their efficacy and product safety record. Excessive heat treatment may, how-ever, cause undesirable protein denaturation, non-enzymatic browning and loss of vitamins and volatile flavor com-pounds. Many US consumers, for example, consider the ‘cooked’ and caramelized flavors of sterilized milk as taste defects[2]. Advances in technology allowed optimization of thermal processing for maximum efficacy against microbial contaminants and minimum deterioration of food quality. High-temperature short-time (HTST) pasteurization and ultra-high temperature (UHT) sterilization, for example, minimize vitamin losses in milk in comparison with batch pasteurization and conventional commercial sterilization, respectively [3]. Products processed by modern thermal technologies, however, still lack the fresh flavor and texture. Non-thermal alternative technologies have been investi-gated intensively in the past 30 years (Table 1)[4–9]. These technologies are named according to the main processing parameter leading to cell inactivation. Food treated with
* Corresponding author. Tel.: +1-614-292-7814; fax: +1-614-292-0218. E-mail address: yousef.1@osu.edu (A.E. Yousef).
www.elsevier.com/locate/micinf
© 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved. PII: S 1 2 8 6 - 4 5 7 9 ( 0 2 ) 0 1 5 5 7 - 5
high-pressure processing (HPP) is exposed to a high hydro-static pressure (up to 1000 MPa) for a few minutes [10]. Pulsed electric field (PEF) treatment is based on the delivery of pulses at high electric field intensity (5-55 kV/cm) for a few milliseconds [11]. Gamma radiations and electron beams generate doses of 2-10 kGy, and are commonly referred to as ionizing radiations [4]. Electron-beam tech-nology is currently developed as a safer alternative to gamma radiation since radioactive isotopes are not used. Ultraviolet (UV) energy is a non-ionizing radiation with germicidal properties at wavelengths in the range of 200-280 nm[8,9].
Alternative technologies are occasionally described as ‘non-thermal’. Food is treated at ambient or refrigeration temperatures and heat generation during the process is not substantial. HPP and gamma radiation are more suitable than other alternative technologies for application in solid foods (Table 1). The lethal agent must penetrate these foods uniformly without degrading the food texture. The current design of PEF treatment chambers does not allow process-ing of solid foods. The shieldprocess-ing effect of solid particles restricts applications of UV radiation and electron beams to the treatment of food surfaces [9]. Some alternative tech-nologies are inherently batch processes, while others are adaptable to continuous applications. HPP was designed primarily for processing of packed food, but new designs were developed recently for continuous processes. PEF is a rapid treatment that is well adapted to continuous
process-ing. Maximal PEF treatment intensity is limited by equip-ment design and food’s ability to withstand dielectric breakdown[12]. Maximal treatment intensity in HPP is also limited by equipment design, particularly when adiabatic heating is to be avoided. Irradiation is a batch process that is easily adapted to continuous applications. When irradi-ated, raw meats and seafood are commonly treated frozen or chilled[4].
3. Efficacy of alternative preservation methods, compared to heat
Most alternative preservation processes achieve the equivalent of pasteurization, but not sterilization[10,13,14]. For illustration purposes, results from different studies will be used to compare alternative preservation methods with a mild heat treatment (Tables 2 and 3) [13–19]. Substantial reductions in the population of Escherichia coli are possible using alternative technologies; these reductions are compa-rable to those achieved by heating at 63 °C for 16 s. HPP requires a comparatively long treatment time. This process may be combined with heat and applied intermittently for elimination of spores[13]. Treatment of food with PEF is a rapid process for inactivating vegetative cells such as
E. coli. The current status of PEF technology does not
enable it to be used to inactivate bacterial spores [14]. Gamma irradiation is effective against vegetative and sporu-lated bacteria [4].
Table 1
Comparison of alternative food-preservation technologies
Process Range of
intensity
Solid food Fluid food Approval Reference
Ionizing radiation 2-10 kGy Raw poultry meat, raw red meat, seafood, spices
Liquid eggs In 41 countries
(http://ccr.ucdavis.edu/irr/inus2.shtml) [4] High pressure processing 100-1000 MPa Ham, seafood Fruit juice, guacamole,
jam, salad dressing, milk
In Japan, north America, Europe [5,6] Pulsed electric field 5-55 kV/cm – Fruit juice, liquid eggs Limited approval in the US (FDA, no
objection letter from 07/07/1995)
[7] Ultraviolet radiation 0.5-20 J/m2 Meat surface, shell egg
surface
Orange juice Approval pending in the US [8,9] .
Table 2
Inactivation of E. coli in milk by heat and alternative preservation technologies
Preservation process Treatment conditions Log count decreased Refs.
Heat 63 °C, 16.2 s 5.9 [15]
γ-Irradiation 10 kGy 7.0 [16]
High-pressure processing 500 MPa, 5 min, 25 °C 5.9 [17]
Pulsed electric field 22.4 kV/cm, 330 µs 4.7 [14]
. Table 3
Inactivation of Bacillus spp. spores by heat and alternative preservation technologies
Technology Treatment conditions Medium Targeted spores Log count decreased Refs.
Heat 140 °C, 3 s Milk Bacillus stearothermophilus 3 [18]
γ-irradiation 12 kGy Frozen yogurt Bacillus cereus 3 [19]
High-pressure processing 2×5 min, 600 MPa, 70 °C Soil medium Bacillus stearothermophilus 3 [13]
Pulsed electric field 22.4 kV/cm, 250 µs Milk Bacillus cereus 0 [14]
The greatest advantage of alternative processes, particu-larly HPP and irradiation, is their efficacy against microor-ganisms at ambient, chilling and freezing temperatures
[4,7,20]. Holding food at mild temperatures favors rapid multiplication of pathogens, spore germination and toxin production. It is safer, therefore, to apply alternative pres-ervation processes at chilling (<5 °C) or hot (45-60 °C) temperatures than at ambient temperatures. Inactivation of
Saccharomyces cerevisiae was not observed between 0 and
25 °C after 10 min at 180 MPa (Fig. 1), probably because cell metabolism was active and cell damage (e.g., mem-brane non-selective permeability) was repaired quickly
[21]; higher pressures were required to kill this psy-chrotrophic microorganism at this temperature range. The greatest inactivation was accomplished when S. cerevisiae was treated at –10, –20 and 48 °C. Treating food with gamma irradiation and HPP at chilling or freezing tempera-ture maintains heat-labile compounds, which contribute to the ‘fresh attributes’ of food. Temperatures ranging from 45 to 70 °C synergistically enhance the efficacy of HPP or PEF against resistant cells. For example, Bacillus spores are fairly resistant to pressures>1200 MPa for several hours at 25 °C, but not to a 600 Mpa oscillatory pressurization at 50–70 °C for 30 min[13]. Heat treatment at 70 °C would be insufficient to kill these spores without pressure.
4. Mechanisms of microbial inactivation
Microorganisms are inactivated when they are exposed to factors that substantially alter their cellular structure or physiological functions. Structural damage includes DNA strand breakage, cell membrane rupture or mechanical damage to cell envelope. Cell functions are altered when key enzymes are inactivated or membrane selectivity is disabled. A preservation technology, e.g. heat, may cause cell death through multiple mechanisms. Limited informa-tion is available about the mechanisms of inactivainforma-tion of microorganisms by alternative preservation technologies. To speed up the implementation of these technologies, research on mechanisms of cell injury and inactivation is urgently needed.
Microbial growth is retarded at pressures in the range of 20-180 MPa; these pressures also inhibit protein synthesis (Fig. 2)[5,22,23]. Loss of cell viability begins at
approxi-mately 180 MPa, and the rate of inactivation increases exponentially as the pressure increases. Lethal high-pressure treatments disrupt membrane integrity and dena-ture proteins[23]. The irreversible denaturation of proteins above 300 MPa corresponds to the range of pressure nec-essary for the inactivation of most vegetative cells and bacteriophages, i.e. protein-coated viruses. Lipid-coated viruses such as the Sindbis virus can resist up to 700 MPa
[6]. Membrane disruption is likely responsible for the changes in morphology observed in HPP-treated cells[5]. The formation of pores in spore coats during treatment at 50-300 MPa may indicate that HPP induces spore germina-tion. No germination was observed at higher pressures, likely because the environmental conditions were poten-tially lethal to germinating spores[6].
Membrane structural or functional damage is generally accepted as the cause of cell death during exposure to high-voltage electric field. Zimmermann[24]suggested that PEF temporarily increases the trans-membrane potential of cells by accumulating compounds of opposite charges in the membrane surroundings. High trans-membrane potential exerts pressure on the cell membrane; this pressure de-creases membrane thickness and ultimately causes pore formation. On the other hand, Tsong [25] introduced the poration theory to explain the mechanism of cell death by PEF. Electroporation in protein channels and lipid domains results in osmotic swelling of the cell and thus membrane weakening until the cell bursts. In both hypotheses, exces-sive pore formation causes irreversible loss of membrane functions. In addition, other researchers suggested different mechanisms for the biocidal action of electricity. Electric discharge in liquid media may generate small amounts of microbicidal agents such as chlorine, free radicals and H2O2 that alter the DNA and cytoplasmic activity during the treatment[26].
Fig. 1. Inactivation of S. cerevisiae after 10 min at 180 Mpa. Adapted from Hashizume et al.[21]. N, count after treatment; N0,initial count.
Fig. 2. Structural and functional changes in microorganisms at different pressures[5,22,23].
Ionizing and UV radiations damage microbial DNA and to a lesser extent denature proteins[8,27]. Potentially lethal DNA lesions are scattered randomly through the cell popu-lation during ionizing and UV radiations. Cells that are unable to repair their radiation-damaged DNA die. Sub-lethally injured cells are often subject to mutations. Ionizing radiations generate hydroxyl radicals from water, which remove hydrogen atoms from the sugar and the bases of the DNA strands. UV energy at 254 nm induces the formation of pyrimidine dimers; this distorts the DNA helix and blocks cell replication. In addition, UV radiation cross-links aro-matic amino acids at their carbon–carbon double bonds. The resulting denaturation of proteins contributes to membrane depolarization and abnormal ionic flow [28]. Irradiation with long-wave UV (320-400 nm) causes the formation of hydroperoxide radicals in the membrane’s unsaturated fatty acids, which induces changes in membrane permeability
[9]. Exposure of shell eggs to UV light (254 nm) at 4350 µW/cm2 for 15 min reduced the aerobic microbial
population by 3 log10 units[8].
5. Kinetics of microbial inactivation
The most widely recognized theories explaining the death behavior of cell populations in response to exposure to lethal factors are the vitalistic and the mechanistic concepts[29]. The former considers permanent phenotypic variations in the degree of resistance of individual cells within a population. A symmetric, but not normal, distribu-tion of the sensitivity of the populadistribu-tion to the treatment is assumed. High treatment intensity is therefore needed to kill the most resistant cells. In contrast, the mechanistic concept considers that each cell of a population has the same degree of resistance, but only a fraction of the population is affected by the lethal agent at a given time. When microorganisms are treated with heat, the logarithm of cell population normally decreases linearly with the treatment time for constant treatment intensity. The resulting straight logarith-mic plots support the mechanistic concept. The vitalistic concept describes a logarithmic inactivation only when process intensity is high compared to the variations in resistance among the cells within the population. Alternative technologies were also believed to inactivate microorgan-isms logarithmically, as found for Staphylococcus aureus pressure-inactivation in milk (Fig. 3). The decimal reduc-tion time (D-value) corresponds to the treatment time required to reduce the microbial population by 90% at constant treatment intensity. The D-value is calculated from the following equation:
D−Value= t2−t1
Log X2−Log X1 (1) with X1 and X2 corresponding to the viable counts after
treatment times t1and t2, respectively.
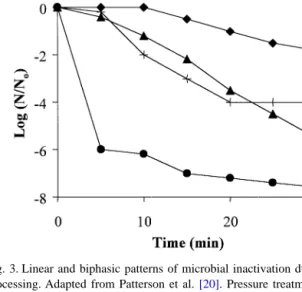
Survivor plots in alternative preservation technologies commonly exhibit a shoulder and/or a tail [20,30](Fig. 3). The initial delay in inactivation of Listeria monocytogenes in milk and S. aureus in poultry meat during HPP for 5 and 10 min, respectively, are typical ‘shoulders’. The intensity of the deleterious agent (e.g. heat) must exceed a threshold value before microbial inactivation happens, which would explain the shoulder. The constant numbers of S. aureus in poultry meat and E. coli in a buffer solution during extended HPP treatment correspond to the tailing effect (Fig. 3). The mechanistic concept considers the shouldering and the tailing effects as an artifact due to factors including sub-lethal injury, cell clumping, and heterogeneous treatment zones. The presence of low electric field regions during the PEF treatment of liquid food has not been clearly estab-lished [30]. On the other hand, presence of resistant sub-populations explains biphasic curves in the vitalistic con-cept. A highly resistant sub-population remains viable over a long period of treatment time, causing a substantial decrease in the efficacy of the process observed at low cell concentration. From a practical point of view, the intensity of the treatment should be high enough to ascertain ‘a virtual absence’ (i.e. undetectable levels) of targeted patho-gens in ready-to-eat food with minimal impact on the product’s quality and production costs.
Numerous models have been developed to fit sigmoid patterns of inactivation; these have been reviewed by Van Gerwen and Zwietering[31]. Inactivation by HPP is initi-ated during the pressure come-up time, i.e. the time needed to raise the pressure to the targeted level. The pressure come-up time depends on the equipment and the headspace in the package[32]. Lethality during come-up time should not be included in constructing survivors’ plots. Kinetics in HPP, therefore, is based on inactivation while the product is at the targeted pressure[20].
Dose–response models are derived from kinetic data to predict the efficacy of the preservation treatment[31]. These
Fig. 3. Linear and biphasic patterns of microbial inactivation during HPP processing. Adapted from Patterson et al.[20]. Pressure treatments: –♦– 375 MPa, L. monocytogenes in milk; –|– 600 MPa, S. aureus in poultry meat; –▲– 600 MPa, S. aureus in milk;–•– 600 MPa, E. coli in 10 mM phosphate saline.
models depict the relationship between treatment intensity and a population resistance parameter. Treatment intensity corresponds to the temperature, pressure, electric field intensity and radiation dose in the case of thermal, high pressure, PEF and radiation treatments, respectively. The
D-value is a commonly used population resistance
param-eter. Heat treatments sufficient to pasteurize or sterilize food are traditionally predicted from thermal death time plots. In linear models, the z-value is an estimate of the treatment intensity necessary to decrease tenfold the D-value:
z−Value= I2−I1
Log D2−Log D1 (2) with I corresponding to the intensity of the preservation treatment.
Dose–response models describing the PEF treatments are based on sigmoid inactivation plots. The survival fraction during the PEF process (X1/X0) at a fixed electric field (E) and a treatment time can be predicted from Fermi’s equation
[33]: X1 X0= 1 1+e
冉
E−Ec k′冊
(3)where k' is the slope of the steepest segment of the survival plot, and Ec, the critical electric field value for 50% survival. For industrial applications, however, both the treatment time (t) and the intensity of the electric field (E) should be considered. The Hulsheger empirical equation takes into account these two process parameters [33]:
X1 X0=
共
t tc兲
−冉
E−Ec k′冊
(4) with tc, the time for inactivation threshold.Hulsheger’s model takes into account the minimum treatment time without loss of viability; therefore, the model describes well the inactivation plots with shoulders. Good fits have been found at high electric strength for both Fermi’s and Hulsheger’s models, probably because the shoulder is reduced at high treatment intensity[33]. Models of Fermi and Hulsheger should be used cautiously, as resistance of microorganisms to PEF increases with cell concentration, which could be due to a shadowing effect, cell clusters or the presence of low electric field regions[7]. To ascertain food safety, an adequate dose-response model must include a safety margin for cell heterogeneity and stress-adaptation mechanisms.
6. Microbial resistance to alternative processes
Bacterial spores are generally the most resistant to inimical processes, followed by positive and Gram-negative bacteria [13]. The higher resistance to HPP and
PEF of Gram-positive, compared to Gram-negative, bacte-ria (Fig. 4) may be linked to the rigidity of the teichoic acids in the peptidoglycan layer of the Gram-positive cell wall. Bacteria of small size and coccoid in shape are generally more resistant to HPP than the large rod-shaped ones[34]. Reduced cell surface area in contact with the environment may limit cell leakage at a given treatment intensity, and thus minimize the effect of the treatment. Resistance to high pressure, however, was greater in S. cerevisiae than bacteria
[34]. This finding indicates that other factors than the shape and size of the cell are involved in cell resistance to alternative preservation treatments. The nature of the cell membrane influences cell resistance to preservation pro-cesses[34]. High membrane fluidity increases the resistance to HPP and low temperatures. Membranes with relatively high fluidity are rich in unsaturated fatty acids. Pressure-resistant cells also have low-diphosphatidylglycerol content
[35]. A specific porin has been associated with increase in pressure resistance[34]. The enhanced resistance of bacte-rial spores to physical stress, solvation and ionization is linked to the protective effect of the membranes and coat layers surrounding the core, the low-water activity in the spore core and the presence of dipicolinic acid[36].
Microorganisms are more likely stressed or injured than killed in food processed by alternative preservation tech-nologies. Adaptation of microorganisms to stress during processing constitutes a potential hazard. Sub-lethal stress induces the expression of cell repair systems (Fig. 5). For instance, exposure of cells to UV induces enzymatic pho-torepair and expression of excision-repair genes that may restore DNA integrity. E. coli that survives UV exposure has an activated htpR gene, i.e. a transcriptional regulator involved in multiple stress resistance. The chaperones GroEL and DnaK, which are under the control of htpR, were isolated post-irradiation[37]. Strains surviving HPP have a similar type of enhanced gene-regulated resistance to both HPP and other stresses[38]. Pressure-induced proteins are
Fig. 4. Lethality of microorganisms after high-pressure processing at 300 MPa (5 °C) for 30 min. Adapted from Arroyo et al.[34]. N, count after treatment; N0, initial count.
produced in a transitory manner, some of which are also induced by heat and cold shocks [39]. The production of high levels of shock proteins is typical of gene-controlled resistance to stress and may contribute to the bacterial stress hardening effect, i.e. the expression of cross-protection mechanisms against multiple types of environmental stresses[40]. Stress-adapted cells are particularly challeng-ing to the food industry; they may survive processes combing several preservation factors (i.e. hurdle technol-ogy). In addition, repetitive exposure of bacterial contami-nants to the preservation process can select for highly resistant mutants[22].
Most alternative preservation technologies affect DNA structure and expression. Reversible DNA supercoiling protects cells against osmotic pressure[6]. Slow replication under sub-optimal growth conditions increases the time allowed for cell repair activity and thus favors recovery of sub-lethally injured microorganisms[41]. The highest sen-sitivity of cell populations is when they are in mid-exponential phase of their growth cycle. This applies to all preservation treatments. Transcriptional regulators such as
rpoS are not expressed at this stage of bacterial growth. The
growth region of yeast cells during budding was found particularly sensitive to PEF[42]. The shift in gene expres-sion and the higher sensitivity of the cell membrane during division therefore contribute to the susceptibility of actively growing cells.
7. Enhancing the efficacy
Adaptation of pathogens to environmental and process-ing stresses constitutes a serious challenge to the food industry[40]. Alternative preservation technologies should inactivate unusually resistant contaminants and prevent or minimize stress adaptation. Improved efficacy was reported with the addition of antimicrobial agents (Fig. 6)[43]. Nisin has a synergistic effect with PEF treatment, and an additive effect with HPP treatment[23,43]. Incorporation of a Nisin
molecule through the membrane may be facilitated during HPP and PEF treatments, which may cause the formation of higher numbers of permanent pores. Ozone contributes to the breakdown of cells during mild PEF treatments [30]. The addition of antimicrobial agents does not, however, eliminate the tailing effect exhibited by the resistant cell sub-population[30,43].
8. Measurement of cell survival
An important issue in food safety is to ascertain that alternative preservation processes kill targeted microorgan-isms rather than merely cause sub-lethal injury or phenotype transition into a resistant dormant state. This minimizes the risk of cell resuscitation and expression of virulence during food storage or after food ingestion [44]. Injured L.
mono-cytogenes, Salmonella typhimurium and E. coli O157:H7
were detected after HPP, when comparing viable counts on non-selective and selective media[45]. The heterogeneity of the cell population after processing is likely due to varia-tions in levels of injury, metabolic activity and viability among the cells. Vital staining with fluorescent dyes showed the presence of viable sub-populations that do not form colonies and are therefore not detected on agar media[34]. It has not been demonstrated whether such viable but non-growing cells are the lineage of scarce injured cells. If this hypothesis were true, an extended lag phase would allow the restoration of the original membrane permeability and metabolic activity[46,47].
Several methods are available to directly assess the extent and nature of cell injury within a microbial popula-tion. These include culturing on selective agar media and cell staining using epifluorescence techniques. Propidium iodide was used to detect cells that lost selective permeabil-ity after HPP [34]. Surface hydrophobicity and ATPase activity may help identify the nature of the membrane damage. The combination of fluorescent dyes in flow cytometric analyses can provide information on different types of cell injury, simultaneously. For instance, ethidium bromide, bis-oxonol and propidium iodide can be used to quantify, in a single experiment, the populations that are reproductively viable, metabolically active and/or with
Fig. 5. Microbial stress, injury, adaptation and resistance to processing.
Fig. 6. Inactivation of Bacillus cereus with 0.06 µg/ml Nisin and PEF treatment at 16.7 kV/cm for 100 µs. Adapted from Pol et al.[43]. N, count after treatment; N0, initial count.
intact polarized membrane [48]. Staining with acridine orange differentiates viable from dead cells, based on their relative proportion of DNA and RNA[48]. The combination of propidium iodide and SYTO 9 in the Live/Dead
Ba-cLightTMsystem gives a similar indication[49]. These dyes
are potentially useful in rapid detection of cell viability and injury during treatment with alternative processes.
9. Microorganisms for efficacy testing
Effective food-preservation processes eliminate hazard-ous pathogens and decrease the loads of spoilage microor-ganisms. The canning industry targets Clostridium
botuli-num and uses heat treatment sufficient to eliminate 12-log of
spores (i.e. a ‘12-D process’). Safe pasteurized milk is ‘virtually free’ of Mycobacterium sp. Challenge and valida-tion studies using food-borne pathogens should not be carried out in the food-processing facility. Surrogate bio-logical indicators have been proposed as alternatives to pathogens in these studies [50]. Surrogate microorganisms should be slightly more resistant than the targeted patho-gens, in order to conservatively estimate the level of pathogens remaining in the treated food. The rapid detection of low levels of pathogens or their corresponding non-pathogenic surrogates may be facilitated when the microor-ganisms are tagged with a selectable marker such as bioluminescence or resistance to antibiotics, provided these mutations are unlikely to be transmitted to other microor-ganisms in the factory [50]. The Clostridium sporogenes spore is the conventional indicator to predict the heat-inactivation of C. botulinum spores. Listeria innocua is a non-pathogenic strain that grows in environments similar to those suitable for L. monocytogenes[50]and this surrogate is suitable for studying efficacy of milk pasteurization. Surrogate organisms for alternative preservation processes are yet to be clearly defined.
10. Conclusion
HPP, PEF, ionizing radiations (gamma and electron beams) and UV light can inactivate food-borne microorgan-isms without substantially heating the food. These technolo-gies are developed to produce safe food with high sensory and nutritional values. The choice of a technique for industrial application depends on food properties and pro-cess design. High pressure and irradiation are the most frequently used alternative technologies, partly because of suitability for solid and liquid food applications. UV radia-tion and electron beams are limited to surface decontami-nation applications. PEF allows the processing of liquid foods rapidly and in a continuous fashion. Antimicrobial agents such as Nisin synergistically enhance the PEF treatment. The combination of HPP or PEF with heat (45-70 ºC) increased their efficacy and reduced bacterial
spore populations. Alternative preservation technologies, however, may not be suitable for food sterilization. Thermal sterilization has remained nevertheless the simplest and most effective method for spore inactivation.
Microbial lethality by HPP and PEF is mainly attributed to changes in the membrane structure and functionality. Nucleic acids are the primary target of ionizing radiations and UV light. Most kinetic studies on alternative technolo-gies reveal a sigmoid inactivation pattern. The tailing effect and cell injury are some of the factors that should be elucidated for improving the efficacy of these new technolo-gies. The role of cell structure, physiology, and gene regulation in microbial resistance to alternative preservation technologies should be investigated.
Acknowledgements
Funds for compiling this article were provided by the Center for Advanced Processing and Packaging Studies and the US National Science Foundation.
References
[1] R. Ahvenainen, New approaches in improving the shelf life of minimally processed fruit and vegetables, Trends Food Sci. Technol. 7 (1996) 179–187.
[2] M.R. Blake, B.C. Weimer, D.J. McMahon, P.A. Savello, Sensory and microbial quality of milk processed for extended shelf life by direct steam injection, J. Food Prot. 58 (1995) 1007–1013. [3] C. Lavigne, J.A. Zee, R.E. Simard, B. Bellveau, Effect of processing
and storage conditions on the fate of vitamins B1, B2, and C and on the shelf-life of goat’s milk, J. Food Sci. 54 (1989) 30–34. [4] J. Farkas, Irradiation as a method for decontaminating food, Int.
J. Food Microbiol. 44 (1998) 189–204.
[5] D.G. Hoover, C. Metrick, A.M. Papineau, D.F. Farkas, D. Knorr, Biological effects of high hydrostatic pressure on food microorgan-isms, Food Technol. 43 (1989) 99–107.
[6] J.P.P.M. Smelt, Recent advances in the microbiology of high pressure processing, Trends Food Sci. Technol. 9 (1998) 152–158. [7] B.L. Qin, U.R. Pothakamury, G.V. Barbosa-Canovas, B.G. Swanson, Nonthermal pasteurization of liquid foods using high-intensity pulsed electric fields, Crit. Rev. Food Sci. Nutr. 36 (1996) 603–627. [8] F.L. Kuo, S.C. Ricke, J.B. Carey, Shell egg sanitation: UV radiation and egg rotation to effectively reduce populations of aerobes, yeasts, and molds, J. Food Prot. 60 (1997) 694–697.
[9] T. Bintsis, E. Litopoulou-Tzanetaki, R.K. Robinson, Existing and potential applications of ultraviolet light in the food industry – a critical review, J. Sci. Food Agric. 80 (2000) 637–645.
[10] R. Gervilla, V. Ferragut, B. Guamis, High pressure inactivation of microorganisms inoculated into ovine milk of different fat contents, J. Dairy Sci. 83 (2000) 674–682.
[11] R. Jeantet, F. Baron, F. Nau, M. Roignant, G. Brule, High intensity pulsed electric fields applied to egg white: effect on Salmonella enteritidis inactivation and protein denaturation, J. Food Prot. 62 (1999) 1381–1386.
[12] Q. Zhang, G.V. Barbosa-Canovas, B.G. Swanson, Engineering aspects of pulsed electric field pasteurization, J. Food Eng. 25 (1995) 261–281.
[13] I. Hayakawa, T. Kanno, M. Tomita, Y. Fujio, Application of high pressure for spore inactivation and protein denaturation, J. Food Sci. 59 (1994) 159–163.
[14] T. Grahl, H. Maerkl, Killing of microorganisms by pulsed electric fields, Appl. Microbiol. Biotechnol. 45 (1996) 148–157.
[15] J.Y. D’Aoust, C.E. Park, R.A. Szabo, E.C. Todd, D.B. Emmons, R.C. McKellar, Thermal inactivation of Campylobacter species, Yersinia enterocolitica, and hemorrhagic Escherichia coli 0157:H7 in fluid milk, J. Dairy Sci. 71 (1988) 3230–3236.
[16] B. Garin-Bastuji, B. Perrin, M.F. Thorel, J.L. Martel, Evaluation of γ-ray irradiation of cows’ colostrum for Brucella abortus, Escheri-chia coli K99, Salmonella dublin and Mycobacterium paratubercu-losis decontamination, Letters Appl. Microbiol. 11 (1990) 163–166. [17] R. Gervilla, X. Felipe, V. Ferragut, B. Guamis, Effect of high hydrostatic pressure on Escherichia coli and Pseudomonas fluore-scens strains in ovine milk, J. Dairy Sci. 80 (1997) 2297–2303. [18] M. Von Konietzko, H. Reuter, Abtötung von Bacillus
stearothermo-philus während der Ultrahocherhitzung von Vollmilch, Milchwis-sensch. 41 (1986) 222–225.
[19] A.E. Hashisaka, J.R. Matches, Y. Batters, F.P. Hungate, F.M. Dong, Effects of gamma irradiation at –78 ºC on microbial populations in dairy products, J. Food Sci. 55 (1990) 1284–1289.
[20] M.F. Patterson, M. Quinn, R. Simpson, A. Gilmour, Sensitivity of vegetative pathogens to high hydrostatic pressure treatment in phosphate-buffered saline and foods, J. Food Prot. 58 (1995) 524–529.
[21] C. Hashizume, K. Kimura, R. Hayashi, Kinetic analysis of yeast inactivation by high pressure treatment at low temperatures, Bio-science 59 (1995) 1455–1458.
[22] K.J.A. Hauben, D.H. Bartlett, C.C.F. Soontjens, K. Cornelis, E.Y. Wuytack, C.W. Michiels, Escherichia coli mutants resistant to inactivation by high hydrostatic pressure, Appl. Environ. Microbiol. 63 (1997) 945–950.
[23] K.J.A. Hauben, E.Y. Wuytack, C.C.F. Soontjens, C.W. Michiels, High-pressure transient sensitization of Escherichia coli to lysozyme and nisin by disruption of outer-membrane permeability, J. Food Prot. 59 (1996) 350–355.
[24] U. Zimmermann, Electrical breakdown, electropermeabilization and electrofusion, Rev. Physiol. Biochem. Pharmacol. 105 (1986) 175–256.
[25] T.Y. Tsong, Electroporation of cell membranes, Biophys. J. 60 (1991) 297–306.
[26] H. Hulsheger, J. Potel, E.G. Niemann, Killing of bacteria with electric pulses of high field strength, Radiat. Environ. Biophys. 20 (1981) 53–65.
[27] L. Lucht, G. Blank, J. Borsa, Recovery of food-borne microorgan-isms from potentially lethal radiation damage, J. Food Prot. 61 (1998) 586–590.
[28] B.E.B. Moseley, in: D.E. Johnston, M.H. Stevenson (Eds.), Food Irradiation and the Chemist, Royal Society of chemistry, Bristol, 1990, pp. 97–108.
[29] O. Cerf, Tailing survival curves of bacterial spores, J. Appl Bacte-riol. 42 (1977) 1–19.
[30] T. Ohshima, K. Sato, H. Terauchi, M. Sato, Physical and chemical modifications of high-voltage pulse sterilization, J. Electrostat. 42 (1997) 159–166.
[31] S.J.C. Van Gerwen, M.H. Zwietering, Growth and inactivation models to be used in quantitative risk assessments, J. Food Prot. 61 (1998) 1541–1549.
[32] B.A. Mertens, High pressure equipment for the food industry, High Pressure Res. 12 (1994) 227–237.
[33] G.V. Barbosa-Canovas, M.M. Gongora-Nieto, U.R. Pothakamury, B.G. Swanson (Eds.), Preservation of Foods with Pulsed Electric Fields, Academic Press, Washington, 1999.
[34] G. Arroyo, P.D. Sanz, G. Prestamo, Response to high-pressure, low-temperature treatment in vegetables: determination of survival rates of microbial populations using flow cytometry and detection of peroxidase activity using confocal microscopy, J. Appl. Microbiol. 86 (1999) 544–556.
[35] N.J. Russell, R.I. Evans, P.F. Steeg, J. Hellemons, A. Verheul, T. Abee, Membranes as a target for stress adaptation, Int. J. Food Microbiol. 28 (1995) 255–261.
[36] W.J. Timson, A.J. Short, Resistance of microorganisms to hydro-static pressure, Biotechnol. Bioeng. 7 (1965) 139–159.
[37] G.C. Walker, Mutagenesis and inducible responses to deoxyribo-nucleic acid damage in Escherichia coli, Microbiol. Rev. 48 (1984) 60–93.
[38] A. Benito, G. Ventoura, M. Casadei, T. Robinson, B. Mackey, Variation in resistance of natural isolates of Escherichia coli O157 to high hydrostatic pressure, mild heat, and other stresses, Appl. Environ. Microbiol. 65 (1999) 1564–1569.
[39] T.J. Welch, A. Farewell, F.C. Neidhardt, D.H. Bartlett, Stress response of Escherichia coli to elevated hydrostatic pressure, J. Bacteriol. 175 (1993) 7170–7177.
[40] Y. Lou, A.E. Yousef, Adaptation to sub-lethal environmental stresses protects Listeria monocytogenes against lethal preservation factors, Appl. Environ. Microbiol. 63 (1997) 1252–1255.
[41] L. Lucht, G. Blank, J. Borsa, Recovery of Escherichia coli from potentially lethal radiation damage: characterization of a recovery phenomenon, J. Food Safety 17 (1997) 261–271.
[42] A.J. Castro, G.V. Barbosa-Canovas, B.G. Swanson, Microbial inac-tivation of foods by pulsed electric fields, J. Food Process. Preserv. 17 (1993) 47–73.
[43] I.E. Pol, H.C. Mastwijk, P.V. Bartels, E.J. Smid, Pulsed-electric field treatment enhances the bactericidal action of Nisin against Bacillus cereus, Appl. Environ. Microbiol. 66 (2000) 428–430.
[44] D. McDougald, S.A. Rice, D. Weichart, S. Kjelleberg, Noncultur-ability: adaptation or debilitation? FEMS Microbiol. Ecol. 25 (1998) 1–9.
[45] N. Kalchayanand, T. Sikes, C.P. Dunne, B. Ray, Hydrostatic pressure and electroporation have increased bactericidal efficacy in combi-nation with bacteriocins, Appl. Environ. Microbiol. 60 (1994) 4174–4177.
[46] G. Bogosian, P.J.L. Morris, J.P. O’Neil, A mixed culture recovery method indicates that enteric bacteria do not enter the viable but non-culturable state, Appl. Environ. Microbiol. 64 (1998) 1736–1742.
[47] T.V. Votyakova, A.S. Kaprelyants, D.B. Kell, Influence of viable cells on the resuscitation of dormant cells in Micrococcus luteus cultures held in an extended stationary phase: the population effect, Appl. Environ. Microbiol. 60 (1994) 3284–3291.
[48] G. Nebe von Caron, P. Stephens, R.A. Badley, Assessment of bacterial viability status by flow cytometry and single cell sorting, J. Appl. Microbiol. 84 (1998) 988–998.
[49] L. Boulos, M. Prevost, B. Barbeau, J. Coallier, R. Desjardins, LIVE/DEAD BacLightt: application of a new rapid staining method for direct enumeration of viable and total bacteria in drinking water, J. Microbiol. Meth. 37 (1999) 77–86.
[50] P.M. Foegeding, N.W. Stanley, Listeria innocua transformed with an antibiotic resistance plasmid as a thermal-resistance indicator for Listeria monocytogenes, J. Food Prot. 54 (1991) 519–523.