DETECTIE VAN PLANTENVIRUSSEN
DOOR
MIDDEL
VAN
NEXT
GENERATION
SEQUENCING
EN
VIRUS ELIMINATIE DOOR
THERMO-THERAPIE BIJ DIOSCOREA SPP.
Aantal woorden: 16.089Anita Mihaylova
Stamnummer: 01600100Promotor: Prof. Dr. Ir. Stefaan Werbrouck
Masterproef voorgelegd voor het behalen van de graad master in de richting Master of Science in de industriële wetenschappen: biochemie
DETECTIE VAN PLANTENVIRUSSEN
DOOR
MIDDEL
VAN
NEXT
GENERATION SEQUENCING EN VIRUS
ELIMINATIE
DOOR
THERMO-THERAPIE BIJ DIOSCOREA SPP.
Aantal woorden: 16.089Anita Mihaylova
Stamnummer: 01600100Promotor: Prof. Dr. Ir. Stefaan Werbrouck
Masterproef voorgelegd voor het behalen van de graad master in de richting Master of Science in de industriële wetenschappen: biochemie
Auteursrechtelijke bescherming
De auteur en promotor geven de toelating deze thesis voor consultatie beschikbaar te stellen en delen van de thesis te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de beperking van het auteursrecht, in het bijzonder met betrekking tot de verplichting van de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze thesis.
25/05/2020
Auteur Promotor
Voorwoord
Deze masterproef werd mogelijk gemaakt dankzij de hulp van Dr. Kris De Jonghe en Yoika Foucart en uiteraard mijn promotor Prof. Stefaan Werbrouck. Ik wens hun hierbij persoonlijk te bedanken voor alle hulp en tijd die ze voor mij hebben vrijgemaakt. Yoika heeft me heel veel bijgebracht over de verschillende technieken die werden gehanteerd. Dankzij de hulp en middelen die door het Instituut voor Landbouw, Visserij en Voedingsonderzoek (ILVO) werden aangeboden konden uitgebreide analyses uitgevoerd worden die hebben bijgedragen tot de verzamelde data.
Preambule
Het doel van deze masterproef was het virusvrij maken van yam cultivars. Hierbij werd voor de detectie van de aanwezige virussen HTS gebruikt en thermotherapie voor het virusvrij maken zelf. De stappen die hiervoor ondernomen moesten worden waren:
1. Het aanmaken van een stockcultuur van alle beschikbare cultivars.
2. Van elk cultivar moest een RNA-extractie uitgevoerd worden. Het RNA zou gebruikt worden als basis voor Illumina sequencing.
3. Een Illumina sequencing en de identificatie van alle aanwezige virussen aan de hand van de bekomen data.
4. Vervolgens zou het RNA ook gebruikt worden voor de aanmaak van cDNA. 5. Het cDNA werd als template gebruikt in de PCR-analyses. Deze werden uitgevoerd
voor het bevestigen van de aanwezigheid van de verschillende virussen, gedetecteerd aan de hand van Illumina sequencing, in de verschillende cultivars.
De aanwezige cultivars dienden vervolgens onderworpen te worden aan thermotherapie. Nadien zouden van de behandelde culturen meristeemculturen worden opgesteld. De meristeemculturen moesten uitgroeien en vermeerderd worden zodat deze nadien opnieuw konden worden getest op de aanwezigheid van de verschillende gedetecteerde virussen. De thermotherapie werd opgestart voor de start van de quarantaineperiode. Een deel van de meristeemculturen werden gestart door Prof. Stefaan Werbrouck die nog toegang had tot het universitair laboratorium. Dankzij dit kon nog data verzameld worden over de levensvatbaarheid van de meristemen. De aanwezigheid van de voordien gedetecteerde virussen diende nog bevestigd te worden bij de culturen verkregen uit de meristemen. Dit was helaas niet mogelijk door de ingevoerde maatregelen ten gevolge van de Covid-19 pandemie. Er werden geen bijkomstige proeven uitgevoerd. De verzamelde data werden als voldoende beschouwd voor het indienen van deze masterproef.
Deze preambule werd in overleg tussen de student en de promotor opgesteld en door beiden goedgekeurd.
Abstract
Yam (Dioscorea spp.) is een van de meest geteelde knolgewassen in Afrika, Azië en Zuid-Amerika. Het gewas wordt vegetatief vermeerderd door de knollen in stukken te snijden en weer te planten. Het voordeel van deze techniek is dat de goede eigenschappen van de moederplant worden doorgegeven. Het nadeel is dat ook bepaalde pathogenen, waaronder virussen, die aanwezig zijn in de moederplant kunnen worden overgedragen.
Virussen zorgen voor een lager rendement en kleinere knollen. In extreme gevallen kunnen ze zelfs zorgen voor een compleet oogstverlies. Daarnaast vereisen ze veel energie van hun gastheer. Het verkrijgen van clean stocks of met andere woorden virusvrije planten kan het oogstrendement verhogen, wat de welvaart van deze landen ten goede komt. Daarnaast kunnen
clean stocks gebruikt worden voor het behoud van germplasma van economisch belangrijke
gewassen zoals yam.
In vitro lijnen van Cubaanse en Nigeriaanse yamgenotypes werden gescreend op de
aanwezigheid van virussen door middel van high throughput sequencing (HTS; ook wel next-generation sequencing of NGS genoemd), namelijk Illumina sequencing en vervolgens bevestigd door PCR. Volgende virussen werden bevestigd bij de beschikbare yam cultivars: badnavirussen (Dioscorea bacilliform virus, Dioscorea bacilliform AL virus, Dioscorea
bacilliform RT virus), potyvirussen (Yam mild mosaic virus), Dioscorea virus A, Dioscorea mosaic-associated virus, Yam chlorotic necrosis virus, Yam necrosis-associated virus, Yam yellow spot mosaic virus en Yam virus Y. De potentiële clean stocks werden bekomen door het
toepassen van in vitro thermotherapie. De groei bij verhoogde temperatuur belet vaak het verspreiden van de virussen tot in de apex van de plant. Het afzonderen van de topmeristemen van deze planten en het opgroeien hiervan kan leiden tot virusvrije culturen. De bekomen culturen uit de meristemen dienen echter opnieuw getest te worden op de aanwezigheid van de voordien gedetecteerde virussen. Enkel op deze manier kunnen de culturen als virusvrij gekenmerkt worden.
Kernwoorden: Next Generation Sequencing, thermotherapie, clean stock, yam,
Abstract
Yam (Dioscorea spp.) is one of the most cultivated tuber crops in Afrika, Asia and South America. It is most often propagated by tearing the tubers and replanting them. The advantage of this technique is that the positive traits of the mother plant are transmitted to the next generation. The downside is that pathogens including viruses, are also transmitted if present in the mother plant.
Viruses can cause a generally lower yield and smaller tubers. In extreme cases, total loss of the cultivar is possible. Viruses also demand a lot of energy from their host. Obtaining clean stocks, meaning virus-free crops, can increase the crop yield, which would be beneficial for the welfare of the countries they are cultivated in. Clean stocks can also be used for the preservation of germplasm from economically important crops like yam.
In vitro lines of Cuban and Nigerian yam genotypes were screened for the presence of viruses using high throughput sequencing (HTS; also referred to as Next Generation Sequencing or NGS), more specifically Illumina sequencing and afterwards confirmed by PCR. Following viruses were confirmed in the available yam cultivars: badnaviruses (Dioscorea bacilliform
virus, Dioscorea bacilliform AL virus, Dioscorea bacilliform RT virus), potyviruses (Yam mild mosaic virus), Dioscorea virus A, Dioscorea mosaic-associated virus, Yam chlorotic necrosis virus, Yam necrosis-associated virus, Yam yellow spot mosaic virus and Yam virus Y. Potential
clean stocks were obtained by thermotherapy of yam plantlets. The growth in higher temperatures often prevents the diffusion of viruses to the apex of the plant. By excising the top meristems of these plantlets and growing them out, virus-free plants could be obtained. The final cultures must be tested again for the presence of the previously present viruses. Only then can the new cultures be labelled as clean.
Inhoudsopgave
Inleiding 11
1 Literatuurstudie 13
1.1 Het belang van yam ... 13
1.2 Invloed van virussen ... 14
1.2.1 Virusdetectietechnieken ... 18
1.2.2 Viruseliminatiemethoden ... 28
2 Experimenteel gedeelte 31 2.1 Vermeerdering van stockculturen ... 31
2.1.1 Doel ... 31 2.1.2 Materiaal en methode ... 31 2.1.3 Resultaten ... 33 2.1.4 Bespreking ... 33 2.1.5 Conclusie ... 33 2.2 Thermotherapie ... 33 2.2.1 Doel ... 33 2.2.2 Materiaal en methode ... 34 2.2.3 Resultaten ... 34 2.2.4 Bespreking ... 34 2.2.5 Conclusie ... 35 2.3 Meristeemculturen ... 35 2.3.1 Doel ... 35 2.3.2 Materiaal en methode ... 36 2.3.3 Resultaten ... 38 2.3.4 Bespreking ... 39 2.3.5 Conclusie ... 40
2.4 Meristeemculturen voor clean stocks ... 41
2.4.1 Doel ... 41
2.4.2 Materiaal en methode ... 41
2.4.4 Bespreking ... 42 2.4.5 Conclusie ... 43 2.5 RNA-extractie voor HTS ... 44 2.5.1 Doel ... 44 2.5.2 Materiaal en methode ... 44 2.5.3 Resultaten ... 45 2.5.4 Bespreking ... 46 2.5.5 Conclusie ... 46 2.6 Sequencing ... 46 2.6.1 Doel ... 46 2.6.2 Materiaal en methode ... 46 2.6.3 Resultaten ... 47 2.6.4 Bespreking ... 48 2.6.5 Conclusie ... 50 2.7 cDNA synthese ... 50 2.7.1 Doel ... 50 2.7.2 Materiaal en methode ... 50
2.8 PCR voor bevestigen van virussen ... 51
2.8.1 Doel ... 51 2.8.2 Materiaal en methode ... 51 2.8.3 Resultaten ... 53 2.8.4 Bespreking ... 59 2.8.5 Conclusie ... 60 3 Algemeen besluit 61 4 Samenvatting 64 Bibliografie 66 Bijlagen i
Lijst met figuren
Figuur 1: Yam knollen op markt (Asiedu & Sartie, 2010). ... 14 Figuur 2: Bladeren van Dioscorea spp. met typische symptomen van yam mosaic disease. a: D. rotundata TDr 99/02674 met mozaïeksymptomen, b: D. rotundata cv. Adaka met
chlorotische verkleuringen en c: mottling, d: D. rotundata TDr 00/00168 met bladvervormingen, e: asymptomatische D. rotundata cv. Adaka. Planten a t.e.m. d testten positief voor Yam Mosaic Virus en alle planten testten positief voor Yam virus Y. (Silva et al., 2019) ... 16
Figuur 3: Werking van ELISA voor virusdetectie (Clark & Adams, 1977) ... 18 Figuur 4: Weergave van de PCR-procedure. Tijdens elke cyclus wordt het dubbelstrengig
template DNA gedenatureerd waarna de primersequenties elk aan de corresponderende streng annealen zodat de synthese van een nieuwe streng kan plaatsvinden m.b.v. de aanwezige DNA-polymerase. Bij elke cyclus verdubbelt de hoeveelheid DNA (Dijkstra & de Jager, 1998). ... 21
Figuur 5: Weergave van Taqman®-principe (R. Mumford et al., 2006) ... 22
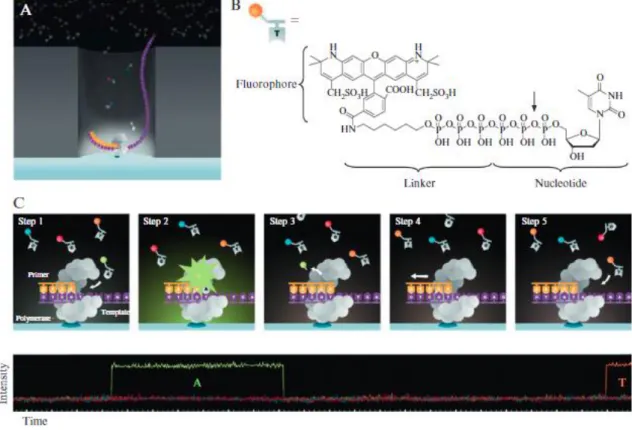
Figuur 6: Illumina sequencing in 4 basisstappen. Stap 1: Willekeurige fragmentatie van DNA
of cDNA en ligatie van adapters aan fragmenten voor vorming van library. Stap 2: Cluster
generation van libraryfragmenten door binding met oligonucleotiden overeenkomstig aan
adaptersequenties en amplificatie. Stap 3: Sequencing door synthese van DNA-strengen m.b.v. fluorofoor gelabelde dNTP’s. Excitatie en detectie van emissie van karakteristieke fluoroforen maken identificatie van de ingebouwde dNTP’s mogelijk. Gedurende elke cyclus wordt één dNTP ingebouwd. Het aantal cyclussen bepaalt bijgevolg de lengte van de reads. Sequencing gebeurt zowel op de forward als de revers streng. Stap 4: Data-analyse van gegenereerde sequenties door koppeling van forward en reverse strengen en mapping ten opzichte van referentiegenomen uit databanken voor identificatie van het organisme. (Illumina, 2017) .... 26
Figuur 7: Principe van SMRTTM DNA-sequencing. (A) In de ZMW nanostructuur arrays wordt telkens 1 polymerase-molecule per compartiment geïmmobiliseerd op de bodem ervan en vervolgens gebonden aan een template DNA-streng. De opbouw van de complementaire streng kan in real-time opgevolgd worden door detectie van de fluorescente labels die ontkoppeld worden tijdens het inbouwen van de dNTP’s. (B) Moleculaire structuur van een fosfogelinkte nucleotide. In dit voorbeeld wordt Alexa Fluor 568-aminohexyltrifosfaat-dTTP weergegeven (Eid et al., 2009). De pijl verwijst naar de plaats waar het fluorofoor wordt losgebonden van de dNTP door de polymerase. (C) SMRTTM DNA-sequencing weergegeven in schematische stappen (bovenaan) en het overeenkomstig fluorescent signaal (onderaan). Stap 1: Het complex van template-DNA, polymerase en primer is omgeven door diffunderende fosfogelinkte nucleotiden. Stap 2: Een gelabeld nucleotide ondergaat een binding met een complementaire base in het template DNA. Terwijl deze wordt ingebouwd in de nieuwe streng ter hoogte van de actieve site van de polymerase wordt een fluorescentiesignaal continu uitgezonden
(gewoonlijk gedurende tienden van milliseconden). Deze puls is detecteerbaar en de identiteit van de gedetecteerde fluorochroom wijst op de base die wordt ingebouwd. Stap 3: De nucleotide werd succesvol ingebouwd in de nieuwe gesynthetiseerde DNA-streng. De thea-bfosfordiester binding tussen het dNTP en de fluorescente label is verbroken en de label wordt vrijgesteld. Stap 4: De polymerase verplaatst zich naar de volgende positie. Stap 5: Het proces wordt herhaald (Korlach et al., 2010). ... 27
Figuur 8: Spectrum in Groeikamer III monochromatisch rood en blauw licht (Philips
production LEDs) ... 32
Figuur 9: Stockculturen. A: nieuwe stekken ‘Belep’ - B: stockcultuur ‘Blanco de Guinea’ - C:
stockcultuur ‘yam 77’. ... 33
Figuur 10: Voorbeeld van in vitro yam cultivar ‘yam 77’ na thermotherapie. A: Verschillende
stekken op medium in groeikamer. B: Close-up van planten in bokaal. C: Close-up van planten op steriel papier vóór dissectie tot meristeemculturen. ... 34
Figuur 12: Meristeemculturen bij opstart van experiment. De samenstelling van de media
wordt weergegeven in Tabel IV (nummer van medium komt overeen). De rijen zijn van boven naar onder respectievelijk: ‘Blanco de Guinea’ met radijswortels, ‘Blanco de Guinea’ zonder radijswortels, ‘yam 77’ met radijswortels en ‘yam 77’ zonder radijswortels. ... 38
Figuur 13: Meristeemculturen na 14 dagen. De samenstelling van de media wordt weergegeven
in Tabel IV (nummer van medium komt overeen). De rijen zijn van links naar rechts respectievelijk: ‘yam 77’ zonder radijswortels, ‘yam 77’ met radijswortels, ‘Blanco de Guinea’ zonder radijswortels, ‘Blanco de Guinea’ met radijswortels, ‘Belep’ zonder radijswortels, ‘Belep’ met radijswortels. ... 39
Figuur 14: Voorbeeld van meristeemculturen geënt op specifiek medium in kleine petriplaat
(Ø55mm). ... 41
Figuur 15: Vorming van scheuten op meristeemculturen. A: Scheutvorming bij cultivar 'yam
77' 2 weken na enting van meristemen. B: Bladvorming bij cultivar 'Belep' 5 weken na enting. ... 43
Figuur 16: Plantjes gevormd door meristeemculturen 4-8 weken na enting op medium. A:
Cultivar ‘Belep’. B: Cultivar ‘yam 77’. C: Cultivar ‘Blanco de Guinea’. ... 43
Figuur 17: Quote Admera voor RNA-sequencing ... 47 Figuur 18: Resultaten PCR-reactie voor Badnavirus. Volgorde van de laantjes: 1. DNA-ladder;
2. yam 18; 3. yam 29; 4. yam 77; 5. yam belep; 6. yam Blanco de Guinea; 7. negatieve controle; 8. duplo yam 18; 9. duplo yam 29; 10. duplo yam 77; 11. duplo yam belep; 12. duplo yam Blanco de Guinea; 13. DNA-ladder ... 53
Figuur 19: Resultaten PCR-reactie voor Yam mild mosaic virus. Volgorde van de laantjes: 1.
negatieve controle; 8. duplo yam 18; 9. duplo yam 29; 10. duplo yam 77; 11. duplo yam belep; 12. duplo yam Blanco de Guinea; 13. DNA-ladder ... 54
Figuur 20: Resultaten PCR-reactie voor Yam virus Y. Volgorde van de laantjes: 1.
DNA-ladder; 2. yam 18; 3. yam 29; 4. yam 77; 5. yam belep; 6. yam Blanco de Guinea; 7. negatieve controle. De duplo stalen werden op een andere gel geladen en gaven hetzelfde resultaat weer, hier niet weergegeven. ... 54
Figuur 21: Resultaten PCR-reactie voor Potyvirus. Volgorde van de laantjes: 1. DNA-ladder;
2. yam 18; 3. yam 29; 4. yam 77; 5. yam belep; 6. yam Blanco de Guinea; 7. negatieve controle; 8. duplo yam 18; 9. duplo yam 29; 10. duplo yam 77; 11. duplo yam belep; 12. duplo yam Blanco de Guinea; 13. DNA-ladder ... 55
Figuur 23: Resultaten PCR-reactie voor Potato yellowing virus (2-7) en Yam yellow spot
mosaic virus (8-13). Volgorde van de laantjes: 1. DNA-ladder; 2. yam 18; 3. yam 29; 4. yam 77; 5. yam belep; 6. staal yam Blanco de Guinea; 7. negatieve controle; 8. yam 18; 9. yam 29; 10. yam 77; 11. yam belep; 12. yam Blanco de Guinea; 13. negatieve controle ... 56
Figuur 22: Resultaten PCR-reactie voor Dioscorea mosaic-associated virus (2-7) en Yam
necrosis-associated virus (8-13). Volgorde van de laantjes: 1. DNA-ladder; 2. yam 18; 3. staal yam 29; 4. yam 77; 5. yam belep; 6. yam Blanco de Guinea; 7. negatieve controle; 8. yam 18; 9. yam 29; 10. yam 77; 11. yam belep; 12. yam Blanco de Guinea; 13. negatieve controle ... 56
Figuur 24: Resultaten PCR-reactie voor Dioscorea virus A (2-7) en Yam chlorotic necrosis
virus (8-13). Volgorde van de laantjes: 1. DNA-ladder; 2. yam 18; 3. yam 29; 4. yam 77; 5. yam belep; 6. yam Blanco de Guinea; 7. negatieve controle; 8. yam 18; 9. yam 29; 10. yam 77; 11. yam belep; 12. yam Blanco de Guinea; 13. negatieve controle ... 57
Figuur 25: Yam cultivars afkomstig van in vitro culturen. A: Cultivar ‘yam 29’ met duidelijk
waarneembare verkleuring van de bladeren aangegeven door de rode kader. B: Cultivar ‘yam 77’ waar geen symptomen op waar te nemen zijn. C: Cultivar ‘yam 18’ met lichte verkleuring op de aangeduide bladeren. D: Cultivar ‘Belep’ met zeer lichte verkleuringen die eventueel te wijten kunnen zijn aan de aanwezige virussen. ... 58
Lijst met tabellen
Tabel I: Meest voorkomende virussen bij Dioscorea spp. (Asala et al., 2012; Eni et al., 2008,
2010; Odu et al., 2004) ... 16
Tabel II: Identificatie van gebruikte yam-soorten. IITA = international institute of agriculture.
... 31
Tabel III: Samenstelling medium voor stockcultuur. ½ MS = Murashige and Skoog basaal
medium met halve concentratie toegevoegde macro- en micronutriënten en vitaminanen; 2iP = isopentenyl adenine (Beyltiens, 2019). ... 32
Tabel IV: Samenstelling van geteste groeimedia voor meristeemculturen van yam. MS =
Murashige and Skoog basaal medium met toegevoegde macro- en micronutriënten en vitaminanen; ½ MS = halve concentratie toegevoegde componenten; 2iP = isopentenyl adenine; BA = 6-benzyladenine; GA3 = gibberellinezuur; IBA = indool-3-boterzuur en NAA =
1-naftaleenazijnzuur. Bronnen: 1: (Salazar et al., 1985)- 2: (Salazar et al., 1985)- 3: (Mantell et al., 1980) - 4: (Filloux et al., 2006) - 5: (Saleil et al., 1990). ... 37
Tabel V: Groei van meristemen op verschillende groeimedia. a : de radijswortels die in het medium werden geplaatst waren omgevormd tot callus begroeid met wortelharen. 1: 1,0mg/L 2iP; 2: 3,0mg/L 2iP + 0,4mg/L NAA; 3: BA + NAA; 4: 2,0g/L actief kool; 5: BA+ GA2 + IBA.
... 38
Tabel VI: Samenstelling van medium gebruikt voor meristeemculturen. De hoeveelheden
worden weergegeven voor 1L medium. ... 41
Tabel VII: Resultaten van meristeemculturen. ... 42 Tabel VIII: Bekomen concentratie door nanodrop absorbantie bepaling na
RNA-extractie. De absorbanieverhouding 260nm:280nm bij de respectievelijke golflengten is een maat voor de contaminatie van het RNA met eiwitten. De gewenste waarde is hiervoor 2,00; een lagere waarde wijst op een contaminatie (T042-TECHNICAL BULLETIN NanoDrop
Spectrophotometers, n.d.). ... 45 Tabel IX: Identificatie virussen na dataverwerking van Illumina sequencing. Indien het genus
ongekend is, wordt de laatste gekende tak van de taxonomie weergegeven in het cursief. ... 48
Tabel X: Samenstelling mastermix voor cDNA-synthese voor 1 staal. ... 51 Tabel XI: Protocol gebruikt voor cDNA-synthese uit RNA-stalen van Dioscorea cultivars . 51 Tabel XII: Samenstelling mastermix voor PCR voor 1 staal. ... 52 Tabel XIII: Gebruikte primer-sequenties voor PCR voor virusdetectie bij beschikbare
yam-cultivars. ... 53
Tabel XIV: Zelfontworpen primers bedoeld voor detectie van de weergegeven virussen door
primersequenties bekomen aan de hand van de PCR Primer Design Tool van Eurofins Genomics (https://www.eurofinsgenomics.eu/en/ecom/tools/pcr-primer-design/) die aan de meeste voorwaarden voldeden, werden geselecteerd. ... 55
Tabel XV: Voorwaarden voor eigen ontworpen primers. De voorwaarden werden nagegaan
aan de hand van de IDT OligoAnalyser (https://eu.idtdna.com/calc/analyzer). a energie vereist
voor het breken van de gevormde structuur ... 56
Tabel XVI: Resultaten van PCR-analysen voor verschillende virussen. De aanwezigheid van
het virus in een bepaalde cultivar wordt weergegeven door ‘x’... 57
Tabel XVII: Protocollen voor PCR-analyses. Indien geen bron wordt vermeld werd de
annealing-temperatuur geschat als het gemiddelde van de smelttemperaturen van de primers verminderd met 5°C. De rest van het verloop is gebaseerd op het standaardprotocol gebruikt aan de ILVO. Bij elke PCR-analyse werden 40 cycli uitgevoerd (denaturatie-annealing-elongatie). Na het laatste cyclus volgde een finale elongatie van 10min bij 72°C. ... ii
Lijst met afkortingen
Afkorting Betekenis
2iP Isopentenyl adenine
BA 6-benzyladenine
CMV Cucumber mosaic virus
cDNA Complementair DNA
DAS ELISA Double Antibody Sandwich Enzyme Linked Immunosorbent Assay
DBALV Dioscorea bacilliform AL virus
DBRTV Dioscorea bacilliform RT virus
DBV Dioscorea bacilliform virus
ddNTP Dideoxyribonucleoside trifosfaten
DMaV Dioscorea mosaic-associated virus
DMoV Dioscorea mottle virus
DNA Desoxyribonucleinezuur
dNTP Deoxyribonucleoside trifosfaten
ELISA Enzyme-linked immunosorbent assay
FRET Fluorescent resonance energy transfer
GA3 Gibberellinezuur
HTS High Throughput sequencing
IBA Indool-3-boterzuur
ILVO Instituut voor landbouw-, visserij- en voedingsonderzoek
LAMP Loop-mediated isothermal amplification
MS Murashige and Skoog
NAA 1-naftaleenazijnzuur
NGS Next generation sequencing
PCR Polymerase chain reaction
PLRV Potato leaf roll virus
PVY Potato virus Y
RNA Ribonucleïnezuur
rpm Rotaties per minuut
RT-PCR Revers transcriptase PCR
UV Ultraviolet
YCNV Yam chlorotic necrosis virus
YMMV Yam mild mosaic virus
YMV Yam mosaic virus
YNaV Yam necrosis-associated virus
YVY Yam virus Y
Inleiding
Yam (Dioscorea spp.) is een van de meest geteelde knolgewassen in Afrika, Azië en Zuid-Amerika. Het gewas wordt vegetatief vermeerderd door de knollen in stukken te snijden en weer te planten. Het voordeel van deze techniek is dat de goede eigenschappen van de moederplant worden doorgegeven. Het nadeel is dat ook bepaalde pathogenen, waaronder virussen, die aanwezig zijn in de moederplant kunnen worden overgedragen.
Virussen zorgen voor een lager rendement en kleinere knollen. In extreme gevallen kunnen ze zelfs zorgen voor een compleet oogstverlies. Daarnaast vereisen ze veel energie van hun gastheer. Het verkrijgen van clean stocks of met andere woorden virusvrije planten kan het oogstrendement verhogen, wat de welvaart van deze landen ten goede komt. Daarnaast kunnen
clean stocks gebruikt worden voor het behoud van germplasma van economisch belangrijke
gewassen zoals yam.
Het doel van deze masterproef was tweeërlei. Enerzijds om na te gaan of er virussen aanwezig waren in 5 verschillende Dioscorea cultivars aan de hand van high throughput sequencing (HTS), meer bepaald Illumina sequencing. Anderzijds om te proberen deze cultivars virusvrij te maken en clean stocks te produceren door middel van in vitro thermotherapie en meristeemculturen.
Deze masterproef is opgebouwd uit een literatuurstudie en een experimenteel gedeelte. In de literatuurstudie wordt nader ingegaan op het belang van yam en de effecten van virussen hierop. Verder wordt er gekeken naar de evolutie van verschillende virusdetectietechnieken. Tot slot wordt besproken op welke manier geïnfecteerde planten virusvrij kunnen gemaakt worden zonder planteigen eigenschappen te verliezen.
De proeven in verband met virusdetectie werden uitgevoerd aan het Laboratorium Virologie van het ILVO van Dr. Kris De Jonghe. Na het uitvoeren van een RNA-extractie werden de stalen gepoold en opgestuurd voor Illumina sequencing. Toen moest de rauwe date geanalyseerd worden met de gepaste software. Vermits de sequencing uitgevoerd werd op een gepoold staal van de verschillende cultivars, dienden de virussen nadien bevestigd te worden bij elk cultivar afzonderlijk. Om dit te verwezenlijken werd van elk RNA-staal cDNA gesynthetiseerd waarna PCR-testen werden uitgevoerd voor de detectie van de verschillende gesequeneerde virussen per cultivar. Bijbehorende resultaten werden besproken en vergeleken met resultaten van gelijkaardige onderzoeken.
De in vitro proeven werden uitgevoerd in het laboratorium voor Toegepaste In Vitro Biotechnologie van Prof. Stefaan Werbrouck. In vitro culturen van vijf yam cultivars werden onderworpen aan thermotherapie. Na deze bijzondere stressvolle behandeling werd erin geslaagd om een aantal meristemen te laten uitgroeien tot een plantje. Opnieuw werden de bekomen resultaten besproken en vergeleken met resultaten van gelijkaardige onderzoeken. Het was de bedoeling was om de resulterende plantjes opnieuw te testen met PCR om na te gaan er virussen geëlimineerd waren. Deze stap is niet meer uitgevoerd omwille van de beperkingen aan de Universiteit van Gent ten gevolge van de Covis-19 pandemie.
Tot slot kan een samenvatting van alle bevindingen en een algemene conclusie teruggevonden worden in deze masterproef. Hierin worden de voornaamste resultaten samengevat en wordt mogelijk vervolgonderzoek besproken.
1
Literatuurstudie
1.1 Het belang van yam
Yam is een knolgewas dat voornamelijk geteeld wordt in Afrika, Azië en een deel van Zuid-Amerika. In West en Centraal Afrika is het de voornaamste voedselbron voor 60 miljoen mensen (Asiedu & Sartie, 2010). Voor de plaatselijke boeren betekent het dan ook een belangrijke bron van inkomsten. Bovendien kan het gebruikt worden in de farmaceutische industrie. Een voorbeeld hiervan is het gebruik van de actieve stof diosgenine, die kan teruggevonden worden in Dioscorea spp., voor de behandeling van neurologische aandoeningen (Cai et al., 2020). Yam wordt daarom als het vierde meest belangrijke knolgewas beschouwd, na de aardappel, de cassave en de zoete aardappel (Sukals et al., 2017). Door de voortdurend toenemende afhankelijkheid van yam in de Afrikaanse landen, wordt de veredeling gestimuleerd om het rendement op te drijven. Hierbij kunnen echter aanwezige virussen worden overgedragen naar de volgende oogst. In sommige gevallen kunnen ook nieuwe virussen of andere pathogenen worden geïntroduceerd door vectoren waardoor verdere oogstverlies kan optreden. Heel wat species worden geteeld zoals: Dioscorea alata, D. bulbifera, D. cayenensis,
D. esculenta, D. opposita-japonica, D. nummularia, D. pentaphylla, D. rotundata en D. trifida.
De meest geteelde hiervan zijn D. rotundata en D. alata.
Yam is een éénjarige plant en de groeicyclus hiervan kan tussen 6 en 12 maanden duren, startende bij de kieming en eindigend bij het afsterven van de plant. Gewoonlijk worden zaaiknollen gebruikt voor het opnieuw aanplanten van het gewas, in plaats van zaden. Zaaiknollen zijn de kleinste knollen van de oogst ofwel in stukken gesneden grote knollen (Asiedu & Sartie, 2010). Zaaiknollen worden verkozen boven zaden omdat deze van de vorige oogst kunnen genomen worden en er op deze manier niet geïnvesteerd dient te worden in zaden. Daarnaast kunnen knollen geselecteerd worden van de planten met de grootste knollen en beste opbrengst om het succes van de volgende oogst enigszins te garanderen. Het grootste nadeel hierbij is dat mogelijke ziektekiemen (virussen, bacteriën e.a.) meegenomen worden naar de volgende oogst. Na het planten van de zaaiknollen en het overbruggen van de rustperiode, worden op korte tijd een uitgebreid wortelsysteem en een jonge plant ontwikkeld. Hierbij ontstaan lange ranken die nog weinig bladeren bevatten en een waslaag hebben, die de plant tegen overmatige verdamping van water beschermt. Deze planten groeien vervolgens uit tot volwassen planten die tenslotte afsterven en knollen in de rustfase achterlaten (Asiedu & Sartie, 2010).
De grote diversiteit aan Dioscorea spp. biedt verscheidene mogelijkheden naar beplanting en groeiperiode toe. De knollen van yam vertonen allen een specifieke dormancy periode die verschilt per cultivar. Dit wil zeggen dat de knollen in een rustfase komen na de oogst, of ze nu bewaard worden of meteen opnieuw geplant.
Indien ze meteen worden geplant, moeten ze de droogteperiode na het groeiseizoen overleven. De rustfase waarin de knollen zich op dat moment bevinden laat dit toe daar de knollen op dat moment droogtetolerant zijn. De knollen worden na twee tot vier maanden opnieuw actief waardoor een nieuwe plant zich kan vormen dankzij de reservestoffen aanwezig in de knol. Bij bewaring bepaalt de duur van de rustfase het moment waarop de knollen opnieuw dienen geplant te worden. Indien de knollen voor consumptie zijn bedoeld en dus niet opnieuw worden geplant, kunnen ze maanden bewaard worden zonder te rotten. De bewaarperiode wordt beïnvloed door de duur van de rustfase van de soort (Asiedu & Sartie, 2010). Daarnaast hebben de verschillende cultivars een verschillende maturatieperiode. Door rekening te houden met deze periode bij het planten van de knollen kunnen de boeren ervoor zorgen dat de oogsten elkaar systematisch opvolgen. Op deze manier worden gedurende het grootste deel van het jaar verse knollen voorzien. De combinatie van een variërende rustfase en maturatieperiode maakt van yam een uitzonderlijk toegankelijk gewas. Een voorbeeld van yamknollen wordt weergegeven Figuur 1.
1.2 Invloed van virussen
Een virus kan gedefinieerd worden als een entiteit waarvan het genoom bestaat uit nucleïnezuren onder de vorm van RNA of DNA dat zich reproduceert in levende cellen. Hierbij wordt gebruik gemaakt van de synthetiserende mechanismen van de gastheercel, daar een virus zichzelf niet kan vermeerderen. De gastheercel richt zich tot de synthese van de specifieke viruspartikels, de virionen, die het genetisch materiaal bevatten. Na synthese worden de virussen samengesteld en vrijgelaten om zich te verspreiden naar de omliggende cellen. Het genoom van virussen is ingekapseld in een beschermende laag aan proteïnen, vaak eiwitmantel genaamd.
Virussen onderscheiden zich van andere intracellulaire obligate parasieten op volgende vlakken (Pennington & Ritchie, 1975):
• Virionen (ofwel viruspartikels) kunnen ofwel RNA ofwel DNA bevatten, nooit beide tegelijk.
• Er wordt gebruik gemaakt van de ribosomen van de gastheercel voor de vermenigvuldiging van virussen.
• Om vermenigvuldiging van de virussen te bekomen, worden eerst de verschillende delen van de virussen apart gesynthetiseerd en vervolgens geassembleerd tot nieuwe viruspartikels. Er vindt dus geen groei door deling plaats.
In de meeste gevallen begint een virus te repliceren na infectie van een vatbare plant. Hierdoor wordt het metabolisme van de cel gewijzigd wat kan zorgen voor biochemische en fysiologische veranderingen. Het abnormaal metabolisme in de verschillende weefsels van de plant kan leiden tot verschillende symptomen. Deze kunnen zichtbaar zijn met het blote oog op de oppervlakte van de plant, of enkel detecteerbaar zijn bij nader onderzoek met een licht- of elektronenmicroscoop. Veranderingen van het metabolisme starten in de primair geïnfecteerde cellen, maar kunnen zich verspreiden naar meerdere weefsels en op deze manier de hele plant inpalmen (Walkey, 1991).
Doordat virussen gebruik maken van de celcomponenten van de geïnfecteerde gastcellen, dragen de gastheercel een zware moleculaire last. De synthese van virionen en de assemblage hiervan vergt veel energie van de cellen. De energie wordt hierbij niet enkel verloren tijdens de synthese van de viruspartikels, maar ook tijdens de productie van de bouwstenen, zoals lipiden en aminozuren. Het rechtstreeks gevolg hiervan is dat de cellen niet meer evenveel kunnen inzetten op de productie van celeigen componenten, wat ervoor zorgt dat de gezondheid van de plant achteruitgaat. Bijgevolg wordt de plant minder resistent tegen het virus en kan de infectie verder woekeren naar andere delen van de plant of kan deze zelfs geïnfecteerd raken door een tweede virus (Walkey, 1991).
Op die manier verstoren virussen de groei van planten. In het geval van voedingsgewassen kunnen ze voor belangrijke opbrengstverliezen zorgen. In ontwikkelingslanden, waar de bevolking sterk afhankelijk is van hun eigen oogst, kan dit nefast zijn. Bij aardappelen bijvoorbeeld kan het opbrengstverlies tot 75% oplopen wanneer het potato leaf-roll virus (PLRV), het potato virus X (PVX) of potato virus Y (PVY) aanwezig zijn in de plant (Al-Taleb, Hassawi, & Abu-Romman, 2011). Het opbrengstverlies verhoogt ook met elk jaar dat een geïnfecteerd gewas opnieuw wordt vermeerderd. Zo werd in de studie van Omer & El-Hassan (1992) een initieel verlies van 20% verhoogd tot 36% bij de volgende vegetatieve cyclus. Sommige virussen induceren zichtbare schade aan het deel van het gewas dat boven de grond groeit, maar andere kunnen aanwezig zijn zonder dat dit met het blote oog zichtbaar is.
Deze laatste zijn een groot gevaar mits ze vaak niet op tijd worden ontdekt en het gewas verder wordt geteeld terwijl de opbrengst hiervan blijft afnemen. Voorbeelden van virale symptomen staan weergegeven in Figuur 2. Verlies aan pigment of necrotische mozaïek verraadt het virus. Door de verminderde mogelijkheid tot fotosynthese ontwikkelen in dit geval de yam planten zich onvoldoende en produceren ze hierbij minder of kleinere knollen.
De meest voorkomende virussen bij yam zijn weergegeven in Tabel I. Deze kunnen zichtbare symptomen induceren als weergegeven in Figuur 2 of asymptomatisch blijven (Umber et al., 2014). Naast deze reeds gekende virussen, zijn ook tal van virussen die momenteel nog niet zijn geclassificeerd.
Tabel I: Meest voorkomende virussen bij Dioscorea spp. (Asala et al., 2012; Eni et al., 2008, 2010; Odu et al., 2004)
Genus Species
Potyvirus Yam mosaic virus (YMV)
Yam mild mosaic virus (YMMV) Dioscorea durnetorum virus (DdV)
Badnavirus Dioscorea alata bacilliform virus (DaBV)
Dioscorea sansibarensis virus (DSBV)
Cucumovirus Cucumber mosaic virus (Cmv)
Dioscorea mottle virus (DMoV)
Algemeen kunnen de symptomen veroorzaakt door een bepaald virus afgeleid worden uit de naam van het virus. Zo refereert de term mosaic naar een patroon op de bladeren of andere delen van de plant waarbij een deel van de cellen geïnfecteerd en verkleurd is en een ander niet. De verkleuringen die optreden zijn te wijten aan het verlies aan chlorofyl in de cellen waardoor deze lichter groen worden of volledig kleurloos. Het patroon van de verkleuring kan sterk variëren afhankelijk van het virus.
Figuur 2: Bladeren van Dioscorea spp. met typische symptomen van yam mosaic disease. a: D. rotundata TDr 99/02674 met mozaïeksymptomen, b: D. rotundata cv. Adaka met chlorotische verkleuringen en c: mottling, d: D. rotundata TDr 00/00168 met bladvervormingen, e: asymptomatische D. rotundata cv. Adaka. Planten a t.e.m. d testten positief voor Yam Mosaic Virus en alle planten testten positief voor Yam virus Y. (Silva et al., 2019)
Indien het voornamelijk om ronde verkleurde vlekken gaat, wordt er vaak naar het virus gerefereerd als mottle, chlorotic flecking, spotting en blotching. Wanneer een volledig blad verkleurd door de aanwezigheid van een virus, wordt hiernaar gerefereerd door de term
chlorotic. De oorzaken van de verkleuring zijn een verlaagde chlorofylproductie of de afbraak
van chloroplasten. Daarnaast wijst dit op interne histologische aandoeningen zoals bijvoorbeeld abnormale veranderingen in de intracellulaire vacuolen of van het palissadeparenchym. Verder kan ook necrose geïnduceerd worden door de aanwezigheid van een plantenvirus. Hierbij worden de cellen in delen van de plant afgedood. Dit kan op enkele plaatsen op de bladeren gebeuren of binnen het transportstelsel van de plant. Indien het transportstelsel wordt aangetast, kan de necrose uitbreiden met de dood van de gehele plant als gevolg. Een verscheidenheid aan andere externe symptomen zoals ringspotting, tumoren, vervorming van de bladeren en stengels, de vorming van abnormale vruchten, zaden of pollen, e.a. kunnen te wijten zijn aan plantenvirussen (Walkey, 1991).
Naast de externe virussymptomen vinden ook enkele interne veranderingen plaats ten gevolge van een virusinfectie. Zo worden de zichtbare symptomen van mozaïek of chlorose veroorzaakt door een storing in de chlorofylproductie van de chloroplasten of de afbraak van deze laatste. Tumoren teruggevonden op geïnfecteerde planten wijzen op een abnormale celdeling veroorzaakt door het virus. Naast deze symptomen die zichtbare gevolgen hebben, kunnen ook andere cytologische of histologische veranderingen plaatsvinden. Een voorbeeld van een cytologisch effect is de verandering van de nucleus van een geïnfecteerde cel. Zo kan de nucleolus worden afgebroken of vergroot tijdens de vermeerdering van het virus in de gastheercel wat de werking van de gastheercel verstoort. Verder kunnen veranderingen van de mitochondriën optreden zoals abnormale membraansystemen of het aggregeren van de mitochondriën. Tot slot vinden ook histologische veranderingen plaats, die meestal leiden tot zichtbare afwijkingen. Vaak is een vermeerdering of vermindering van het aantal cellen een gevolg van de afwijking. Een voorbeeld van vermindering van cellen is de vervanging van xyleem- en floëemcellen door parenchymcellen door het apple stem grooving virus. Hierbij worden zichtbare groeven gecreëerd in de stam van de boom. Een vermeerdering van cellen wordt geïllustreerd door de de swollen shoot disease van cacao waarbij een abnormale deling van het cambium leidt tot verhoogde hoeveelheden xyleem waardoor de scheuten van de plant opzwellen (Walkey, 1991).
Tot slot kunnen ook inclusielichamen ontstaan in de nucleus of het cytoplasma van geïnfecteerde cellen. Deze inclusielichamen zijn het gevolg van de vermeerdering van het virus in de plantencellen. Het zijn namelijk samengeclusterde viruspartikels of overschotten van virale proteïnen die niet gebruikt werden tijdens het samenstellen van de viruspartikels. Hun vorm en structuur is afhankelijk van het virus dat ze geïnduceerde (Walkey, 1991).
Bij latente infectie vertonen de geïnfecteerde planten geen symptomen van de virusinfectie. Echter blijven de virussen gerepliceerd worden binnenin de gastheercellen. Dit heeft als gevolg dat, in geval van een gewas, de opbrengst van de plant afneemt ten gevolge van de infectie. Dit wordt echter pas gedetecteerd na vergelijking met de opbrengst van een virusvrije plant (Walkey, 1991). Zo’n latente infectie wordt meestal onbewust doorgekweekt door boeren die gebruik maken van vegetatieve vermeerdering, zoals het geval is bij yam.
1.2.1 Virusdetectietechnieken
De detectie van virussen is niet vanzelfsprekend. Om de aanwezigheid van virussen na te gaan bij bepaalde gewassen wordt vaak gebruik gemaakt van de polymerase chain reaction (PCR) of een enzyme-linked immunosorbent assay (ELISA) (Boonham et al., 2014). Beide technieken kunnen gecombineerd worden om bekomen resultaten te vergelijken en te bevestigen. De voornaamste voordelen van deze zijn dat ze beide een standaardprocedure zijn geworden in de sector en ze zeer reproductief en repetitief zijn. Ze kunnen met andere woorden gemakkelijk meermaals op reproductieve wijze uitgevoerd worden. Dit maakt het vergelijken van bekomen resultaten zeer vanzelfsprekend. Daarnaast zijn deze technieken eenvoudig in uitvoering dankzij de beschikbare technologieën en kunnen ze snel aangeleerd worden aan onervaren laboranten.
Voor testen op het veld zelf, waar geen laboratorium gevestigd is, kan gebruik gemaakt worden van de loop-mediated isothermal amplification (LAMP) techniek. Verder kunnen technieken aangewend worden die de detectie van meerdere virussen tegelijkertijd mogelijk maken zoals bijvoorbeeld Luninex bead arrays. Tenslotte kan ook HTS aangewend worden, een techniek waarbij het volledig genoom kan gesequeneerd worden. HTS wordt vaak gebruikt als screeningstechniek. Voorbeelden van HTS zijn Illumina sequencing en PacBio sequencing.
ELISA
De doorbraak in de diagnose van virussen bij planten gebeurde door Clark & Adams in 1977 dankzij hun publicatie over een ELISA-methode voor de detectie van Plum pox virus (PPV) en
Arabis mosaic virus (ArMV). De detectie van virussen gebeurde
voordien door middel van herkenning van de symptomen door ervaren deskundigen. Er waren reeds immuno-gerelateerde werkwijzen maar deze waren zeer omslachtig en tijdrovend. De introductie van de ELISA-methode was een ware revolutie voor de virusdetectie bij planten (Boonham et al., 2014).
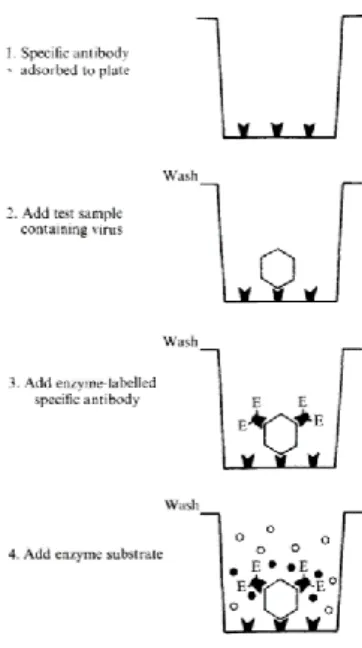
Deze ELISA werkt als volgt (Figuur 3) (Clark & Adams, 1977):
1. Een antilichaam (IgG) specifiek voor het te detecteren virus wordt gecoat op een ELISA-plaat.
2. De niet-gebonden antilichamen worden door middel van een wasstap verwijderd. 3. Een staal met het virus van interesse (antigeen) wordt aan de plaat toegevoegd. Het
antigeen bindt aan het antilichaam en de plaat wordt vervolgens gewassen.
4. Geconjugeerde antilichamen met merker worden aan de plaat toegevoegd. Deze merker is een enzym dat later gebruikt wordt voor de detectie. Deze antilichamen (IgA) binden ook aan het virus en vormen op deze manier een sandwich, als het ware.
5. Opnieuw wordt een wasstap uitgevoerd om overtollig niet gebonden reagens weg te wassen.
6. Een enzym-specifiek substraat wordt aan de plaat toegevoegd zodat een enzymatische reactie, meer bepaald een hydrolyse, kan plaatsvinden. Deze reactie zorgt voor de vorming van een kleurcomponent dat spectrofotometrisch gedetecteerd kan worden. De concentratie van het bekomen product kan gelinkt worden aan de concentratie aan virus aanwezig in het staal.
ELISA is sindsdien de meest gebruikte methode geworden voor de detectie van virussen in gewassen. De belangrijkste voordelen van deze techniek is de robuustheid ervan, het gebruiksgemak, de kosteneffectiviteit en de mogelijkheid voor opschaling om meerdere stalen simultaan te testen. Echter zijn er ook nadelen aan deze methode, namelijk de nood aan antisera van zeer zuivere kwaliteit, die niet evident aan te maken zijn en duur in aanmaak. De productie ervan vergt veel tijd en kennis en is hierdoor enkel mogelijk in gespecialiseerde laboratoria. Verder is ELISA niet gevoelig genoeg om een verschil binnen species te kunnen onderscheiden. Tot slot kan met ELISA telkens voor slechts één virus gescreend worden. Indien er binnen eenzelfde gewas voor verschillende virussen dient gescreend te worden, zal ELISA zeer tijdrovend zijn en hierdoor minder geschikt. Doch blijft ELISA een van de meest gebruikte technieken voor virusdetectie voor het certificeren van gewassen, voor sanering en quarantaine waarbij het belangrijk is dat de virussen op een betrouwbare manier worden gedetecteerd (Boonham et al., 2014).
PCR
De intrede van de PCR-reactie als screeningsmethode dateert sinds midden jaren ’80. Desondanks de vele publicaties en het uitvoerig onderzoek in het hierop volgend decennium werd deze methode niet op grote schaal gebruikt in laboratoria voor de detectie van plantenvirussen. De grote gevoeligheid van deze methode, die eerst als een groot voordeel werd beschouwd, werd al snel het grootste nadeel. Het risico op cross-contaminatie van stalen was namelijk te groot en gaf aanleiding tot verkeerde interpretaties (Boonham et al., 2014; Mumford et al., 2006). Niet de specificiteit of gevoeligheid van de methode moesten verder bestudeerd worden, maar de praktische toepassing ervan.
Voor detectie van plantenvirussen met een RNA-genoom wordt PCR in combinatie met de reverse transcriptie van het RNA (RT-PCR) het vaakst toegepast (López et al., 2003). De RNA-template wordt aan de hand van een revers transcriptase-enzym geconverteerd tot complementair DNA (cDNA) vóór de start van de PCR en dient vervolgens als template voor de PCR (Hughes & Odu, 2003). De enkelvoudige cDNA-streng wordt in de eerste cycli geamplificeerd waardoor een dubbelstrengige DNA-template bekomen wordt voor verdere amplificatie tijdens een standaard PCR-procedure hieronder beschreven. Naast het template DNA is ook een mix van de 4 deoxyribonucleoside trifosfaten A, T, C en G (dNTP’s) noodzakelijk alsook een DNA-polymerase en 2 primersequenties.
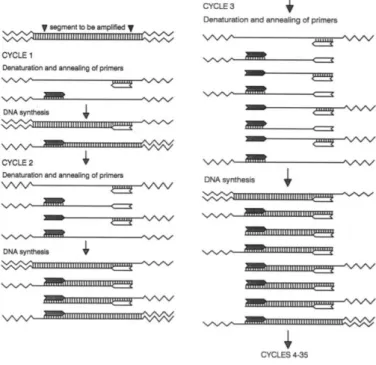
Dankzij PCR kan een DNA-sequentie die zich tussen twee gekende nucleïnezuursequenties bevindt, geamplificeerd worden. De te ampliciferen sequentie wordt geflankeerd tussen twee oligonucleotiden die dienen als primers voor een opeenvolging van reacties gekatalyseerd door DNA-polymerase. Algemeen wordt de PCR opgebouwd uit een initiële denaturatie van het DNA gedurende gemiddeld 3 minuten bij ±95°C, een herhaling van gemiddeld 35 cycli en een finale elongatie van het gesynthetiseerd DNA gedurende 10 minuten bij ±72°C. Tijdens elke cyclus wordt de concentratie aan DNA verdubbeld, anders gezegd geamplificeerd. De cycli bestaan elk uit 3 basisstappen, weergegeven in Figuur 4:
1. De denaturatie van het dubbelstrengig template DNA onder invloed van een verhoogde temperatuur, nl. ±95°C, gedurende gemiddeld 30sec. Hierbij komen de twee DNA-strengen van elkaar los door de breking van waterstofbindingen.
2. De volgende stap is de annealing stap. De annealingtemperatuur is afhankelijk van de smelttemperaturen van de gebruikte primers. Tijdens deze stap hybridiseren de primers aan het template-DNA om zo de basis te vormen van de nieuwe DNA-streng.
3. Tenslotte volgt de elongatie-stap waarin het DNA-polymerase een nieuwe streng synthetiseert op basis van de template-streng.
De gebruikte primers hiervoor zijn gebaseerd op de sequenties van de te detecteren virussen. Deze kunnen achterhaald worden aan de hand van gekende databanken. Algemeen dienen de primers uit 18-25 nucleotiden te bestaan en een smelttemperatuur van 55-65°C te hebben. De targetsequentie die geamplificeerd wordt, heeft bij voorkeur een lengte van 200-500pb (Dijkstra & de Jager, 1998).
Figuur 4: Weergave van de PCR-procedure. Tijdens elke cyclus wordt het dubbelstrengig template DNA gedenatureerd waarna de primersequenties elk aan de corresponderende streng annealen zodat de synthese van een nieuwe streng kan plaatsvinden m.b.v. de aanwezige DNA-polymerase. Bij elke cyclus verdubbelt de hoeveelheid DNA (Dijkstra & de Jager, 1998).
Na amplificatie worden de bekomen sequenties gevisualiseerd aan de hand van agarose gelelektroforese. Hierbij wordt het DNA geladen op een 1,5% agarosegel. Vervolgens wordt de elektroforese uitgevoerd waardoor het DNA wordt gescheiden op basis van grootte op de gel. De elekroforese houdt in dat een zekere spanning wordt gelegd over de gel, bijvoorbeeld 100V, voor een zekere tijd, bijvoorbeeld 40min. Voor het laden van de gel wordt het DNA gekleurd aan de hand van een merker zoals bijvoorbeeld ethidiumbromide zodat het achteraf kan gevisualiseerd worden op de agarosegel aan de hand van bijvoorbeeld UV-licht (Dijkstra & de Jager, 1998). Naast het geamplificeerd DNA kan ook een DNA-ladder geladen worden op de gel. Deze is samengesteld uit verschillende DNA-sequenties met gekende grootte. Door de plaats van de verschillende sequenties van de ladder te vergelijken met de plaats van de sequenties van de amplicons kan de grootte van deze laatste bepaald worden. Indien de grootte van de geamplificeerde sequentie gekend is, kan vervolgens nagegaan worden of de correcte sequentie werd bekomen. Om een mismatch met de plant zelf uit te sluiten, kan de sequentie van het bekomen amplicon bevestigd worden aan de hand van bijvoorbeeld Sanger sequencing.
Real-time PCR
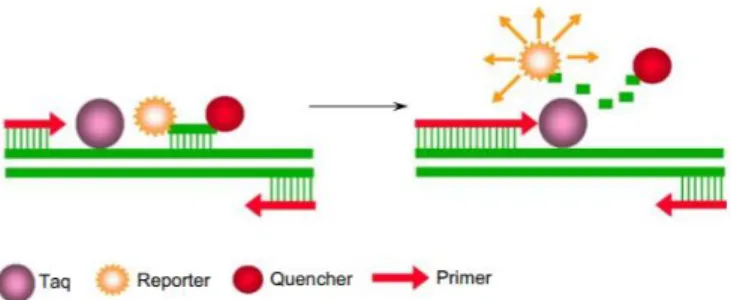
De introductie van het TaqMan®-principe, weergegeven in Figuur 5, maakte van de PCR-methode een van de meest gebruikte PCR-methodes voor virusdetectie in planten. Het werkingsprincipe hiervan is als volgt (R. Mumford et al., 2006):
1. Een probe, bestaande uit een oligonucleotide gelabeld aan beide uiteinden met respectievelijk een reporter en een quencher sequentie, bindt aan een target-sequentie tussenin de twee primer bindingssequenties. De fluorescentie uitgezonden door de reporter wordt opgenomen door de quencher en er wordt geen signaal uitgezonden. Dit wordt beschreven als de fluorescent resonance energy transfer (FRET).
2. Gedurende de amplificatie wordt de probe gesplitst door de 5’-3’endonucleaseactiviteit van het Taq-polymerase. Hierbij wordt de afstand tussen de reporter en de quencher groter waardoor er geen FRET meer plaatsvindt en het fluorescentiesignaal uitgezonden door de reporter spectrofotometrisch gemeten kan worden. De sterkte van dit signaal is recht evenredig met de concentratie aan gevormd amplicon. Wanneer het PCR-toestel uitgerust is met een absorbantiemeter, kan een real-time PCR uitgevoerd worden.
Het voornaamste voordeel van de real-time PCR ten opzichte van de standaard PCR is dat er de reactie meteen kan opgevolgd worden en een kwantificatie van het viraal DNA of RNA kan uitgevoerd worden. Daarnaast vereist real-time PCR geen gel-elektroforese voor het visualiseren van de resultaten. Er zijn bijgevolg ook geen extra manipulaties van het staal nodig na reactie. Verder heeft real-time PCR ook enkele voordelen ten opzichte een ELISA. Ten eerste vereist de PCR-reactie minder voorbereidend werk omdat er geen antiserum dient aangemaakt te worden. Er zijn weliswaar primers en een probe vereist hiervoor, maar de aanmaak hiervan is eenvoudiger. Verder is het uitvoeren van de PCR-reactie minder complex. Een minder gespecialiseerd laboratorium kan deze methode hierdoor gemakkelijker implementeren. Ten slotte is de kost van een PCR-run lager dan deze van een ELISA. De gebruikte reagentia zijn duurder in aankoop, maar de hogere kost voor de aanmaak van antisera voor de ELISA weegt niet op tegen de lagere kost voor de reagentia van de PCR.
LAMP
De introductie van loop-mediated isothermal amplification (LAMP) als virusdetectiemethode maakte het mogelijk om voortaan stalen on-site te analysen (Boonham et al., 2014). Het gemak van deze methode is dat er geen toestel vereist is dat een complex temperatuurprogramma moet doorlopen. Alle stappen kunnen isothermisch uitgevoerd worden waardoor dit kan plaatsvinden in een warmwaterbad of incubator. Daarnaast kan de aan- of afwezigheid van het target DNA of RNA bepaald worden aan de hand van een eenvoudige fluorescentiemeting of een turbiditeitsmeting waardoor een gel-elektroforese overbodig wordt (Yan et al., 2019). Voor een fluorescentiemeting dienen intercalerende fluorochromen, zoals bijvoorbeeld SYBR Green, aan het reactiemengsel toegevoegd te worden. Dit maakt van deze detectiemethode een zeer goedkoop maar even gevoelig alternatief voor PCR.
Het principe van LAMP is gebaseerd op het gebruik van vier primers voor zes target sequenties. De nieuwgevormde strengen worden van de template strengen verwijderd door middel van
stand-displacement. Deze werking is eigen aan de gebruikte polymerase, die de gebonden
streng afduwt tijdens de elongatie van de template-streng. Bijgevolg is geen denaturatie van het DNA vereist voor amplificatie, wat wil zeggen dat de temperatuur ongewijzigd blijft en de reactie isothermisch kan blijven doorgaan bij een temperatuur van ±65°C (Notomi et al., 2015). Detectie van het DNA kan reeds plaatsvinden na 5 minuten en de volledige amplificatie neemt gemiddeld 30 minuten in beslag. In het geval van amplificatie van RNA dient reverse transcriptase aan de mix toegevoegd te worden zodat het dan gevormde cDNA als template voor de reactie kan dienen.
HTS
Bovenstaande methoden worden standaard gebruikt voor de detectie van een beperkt aantal virussen per staal per analyse. Wanneer screening van een bepaald staal wordt vereist, zou de informatie bekomen a.d.h.v. deze methoden beperkt zijn tot de doelgericht gezochte virussen. Virussen waar niet specifiek op wordt getest, zullen hierbij ongedetecteerd blijven. Daarnaast kunnen deze enkel toegepast worden indien de sequentie van het te detecteren virus reeds gekend is. Een recente techniek voor virusdetectie waarmee vele virussen van verschillende types tegelijk kunnen worden geïdentificeerd is high throughput sequencing (HTS). HTS kan chronologisch gezien opgedeeld worden in drie generaties. De eerste generatie sequencing is Sanger sequencing, hieronder verder besproken. Nadien kwam een tweede generatie sequencing op, toen Next Generation Sequencing genaamd, waartoe o.a. Illumina sequencing behoort. Tot slot werden real-time long read sequencing-technieken ontwikkeld in een derde generatie HTS zoals o.a. Pacific Bioscience (PacBio) sequencing.
Sanger Sequencing
Sanger sequencing is een methode gebruikt voor het bepalen van de nucleotidensequentie van DNA of RNA. Hierbij wordt gebruik gemaakt van 2’,3’-dideoxy analogen van normale dNTP’s. Deze analogen werken als inhibitoren van het DNA-polymerase en zorgen ervoor dat de translatie wordt beëindigd.
Er wordt gestart met het aanmaken van 4 stalen met het template DNA waarvan de sequentie dient bepaald te worden. Aan elk staal worden normale dNTP’s toegevoegd samen met één van de vier inhibitoren (ddNTP). Er wordt een DNA-polymerase hieraan toegevoegd en de stalen worden geïncubeerd zodat elongatie van het DNA kan plaatsvinden. Tijdens de incubatie worden fragmenten van verschillende groottes bekomen door het willekeurig inbrengen van een ddNTP in de nieuwe DNA-streng. Door uitzetting op de agarosel kan vervolgens de nucleotidesequentie van het DNA worden afgelezen. De bekomen sequentie kan gevalideerd worden door gebruik van primers op de complementaire streng. (Sanger et al., 1977).
Ontwikkelingen binnen de fluorescentie-technologieën, de enzymologie en de capillaire elektroforese technieken o.a. hebben geautomatiseerde Sanger sequencing mogelijk gemaakt. Hierbij kunnen fluorescente tags geplaatst worden op de ddNTP’s en kan de nucleotidensequentie achterhaald worden aan de hand van fluorescentiemetingen. Dit verkort de analysetijd drastisch en verhoogt de accuraatheid van de data-analyse na sequencing. Door de verhoudingen aan ddNTP/dNTP te laten variëren, kan de lengte van de bekomen reads ook voor een deel bepaald worden (Metzker, 2005).
Illumina sequencing
Illumina sequencing maakt gebruik van dezelfde basisprincipes als Sanger sequencing. Hierbij wordt ook gebruik gemaakt van een DNA-polymerase die een nieuwe streng synthetiseert gebaseerd op een template DNA-streng door het inbouwen van gelabelde dNTP’s. Gedurende elke cyclus kunnen de ingebouwde nucleotiden geïdentificeerd worden door de emissie van een fluorescent signaal. Het voornaamste verschil met traditionele sequencing is dat er gebruik gemaakt wordt van miljoenen DNA-fragmenten die op een parallelle manier gesequeneerd worden daar waar oorspronkelijk slechts één DNA-fragment werd onderzocht. Illumina seuqencing heeft als bijkomende voordelen een zeer hoge accuraatheid en een hoge opbrengst van foutloze reads (Illumina, 2017).
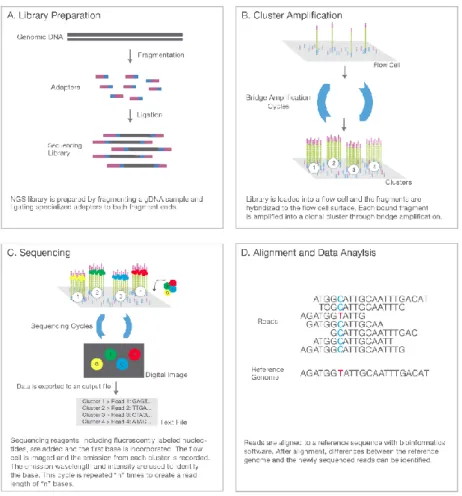
Illumina sequencing-by-synthesis gebeurt volgens vier basisstappen, (Figuur 6):
1. Library preparation: De library voor sequencing wordt klaargemaakt door willekeurige fragmentatie het DNA of cDNA staal dat geanalyseerd dient te worden. Vervolgens worden de 5’ en 3’ adapters geligeerd aan de DNA-templates. De adapters zijn opgebouwd uit een bindingssite voor de sequencing, een indexregio en een deel complementair aan het de oligonucleotiden van de gebruikte flowcellen. Tenslotte worden de adapter-DNA-fragmenten geamplificeerd d.m.v. een PCR-reactie en gezuiverd door gebruik van een gel. 2. Cluster generation: Clustering is de isotherme amplificatie van de DNA-fragmenten. De flowcel hiervoor gebruikt is een glazen plaat met laantjes. Elk laatje is gecoat met een spreekwoordelijk veld aan oligonucleotiden. Er zijn twee soorten oligonucleotiden aanwezig, één voor elk uiteinde van de fragmenten. Tijdens de cluster generation, worden fragmenten gebonden aan één van de twee oligo’s aanwezig in de flowcel. Een polymerase zorgt vervolgens voor de elongatie van de streng complementair aan het template-fragment. Een denaturatiestap zorgt voor het loskoppelen van de template en de gesynthetiseerde streng. Hierna volgt een proces van bridge amplification waarbij de gesynthetiseerde strengen buigen en hybridiseren op een naburig oligonucleotide overeenkomstig met de adapterregio op het vrije uiteinde van de streng.
Een polymerase zorgt opnieuw voor de synthese van een nieuwe streng en de twee strengen worden losgekoppeld in een volgende denaturatiestap. Dit proces wordt herhaald tot alle gebonden oligonucleotiden verlengd werden tot nieuwe gesynthetiseerde strengen, wat resulteert in miljoenen kopieën van alle oorspronkelijke fragmenten. Tenslotte worden alle reverse strengen ontkoppeld zodat enkel de forward strengen gebonden blijven op de flow-cel en de 3’ uiteinden worden geblokkeerd om priming hierop te vermijden.
3. Sequencing: Een eerste primer wordt gebonden aan de aanwezige strengen op de overeenkomstige adapterregio’s en de sequencing van de forward strengen wordt gestart. Gedurende elke cyclus wordt één fluorescent gelabeld nucleotide gebonden aan de nieuwe DNA-streng, complementair aan het template-DNA. Vervolgens worden de clusters geëxciteerd en treedt er emissie op van de fluoroforen overeenkomstig met de ingebouwde nucleotide. Binnen een cluster worden overeenkomstige strengen tegelijkertijd gelezen. Dit proces wordt omschreven als sequencing-by-synthesis. De lengte van de bekomen reads wordt bepaald door het aantal cyclussen.
Na de eerste read, worden de producten weggewassen waarna de volgende primer (index 1) aan de template wordt gebonden en een nieuwe read op een gelijkaardige manier wordt gegenereerd. Hierna wordt de sequentie die het 3’-uiteinde blokkeerde verwijderd waarna de templates opnieuw buigen en binden aan de oligonucleotiden die overeenkomen met de nu vrije uiteinden. Index 2 wordt op dezelfde manier als index 1 afgeschreven. Een polymerase zorgt voor de synthese van de reverse streng waarna de stengen losgekoppeld worden en de forward streng wordt verwijderd. Een tweede sequencing-proces vindt plaats. 4. Data-analyse: De data die gegenereerd wordt door de sequencing bestaat uit miljoenen
read-sequenties. Forward en reverse sequenties worden gekoppeld en creëren zogenaamde contiguous sequences. Deze sequenties worden gealigneerd tegenover referentiegenomen
uit databanken om op deze manier het organisme te kunnen identificeren.
Figuur 6: Illumina sequencing in 4 basisstappen. Stap 1: Willekeurige fragmentatie van DNA of cDNA en ligatie van adapters aan fragmenten voor vorming van library. Stap 2: Cluster generation van libraryfragmenten door binding met oligonucleotiden overeenkomstig aan adaptersequenties en amplificatie. Stap 3: Sequencing door synthese van DNA-strengen m.b.v. fluorofoor gelabelde dNTP’s. Excitatie en detectie van emissie van karakteristieke fluoroforen maken identificatie van de ingebouwde dNTP’s mogelijk. Gedurende elke cyclus wordt één dNTP ingebouwd. Het aantal cyclussen bepaalt bijgevolg de lengte van de reads. Sequencing gebeurt zowel op de forward als de revers streng. Stap 4: Data-analyse van gegenereerde sequenties door koppeling van forward en reverse strengen en mapping ten opzichte van referentiegenomen uit databanken voor identificatie van het organisme. (Illumina, 2017)
PacBio sequencing
PacBio sequencing genereert sequencing data door een enkele DNA-polymerase molecule die continue template-gestuurde DNA-synthese uitvoert (Eid et al., 2009). De gebruikte polymerase bouwt hierbij fluorescent gelabelde dNTP’s in. De label is eigen aan het dNTP en is zodanig gelinkt dat de label ontkoppeld wordt tijdens het inbouwen van de dNTP en de DNA-synthese niet onderbroken wordt. Tijdens het loskoppelen van de label door de polymerase ontstaat een kortstondig en intens fluorescent signaal. Door meting van dit signaal, kan de nucleotidensequentie van de gesynthetiseerde DNA-streng bepaald worden.
Figuur 7: Principe van SMRTTM DNA-sequencing. (A) In de ZMW nanostructuur arrays wordt telkens 1 polymerase-molecule
per compartiment geïmmobiliseerd op de bodem ervan en vervolgens gebonden aan een template DNA-streng. De opbouw van de complementaire streng kan in real-time opgevolgd worden door detectie van de fluorescente labels die ontkoppeld worden tijdens het inbouwen van de dNTP’s. (B) Moleculaire structuur van een fosfogelinkte nucleotide. In dit voorbeeld wordt Alexa Fluor 568-aminohexyltrifosfaat-dTTP weergegeven (Eid et al., 2009). De pijl verwijst naar de plaats waar het fluorofoor wordt losgebonden van de dNTP door de polymerase. (C) SMRTTM DNA-sequencing weergegeven in schematische
stappen (bovenaan) en het overeenkomstig fluorescent signaal (onderaan). Stap 1: Het complex van template-DNA, polymerase en primer is omgeven door diffunderende fosfogelinkte nucleotiden. Stap 2: Een gelabeld nucleotide ondergaat een binding met een complementaire base in het template DNA. Terwijl deze wordt ingebouwd in de nieuwe streng ter hoogte van de actieve site van de polymerase wordt een fluorescentiesignaal continu uitgezonden (gewoonlijk gedurende tienden van milliseconden). Deze puls is detecteerbaar en de identiteit van de gedetecteerde fluorochroom wijst op de base die wordt ingebouwd. Stap 3: De nucleotide werd succesvol ingebouwd in de nieuwe gesynthetiseerde DNA-streng. De thea-bfosfordiester binding tussen het dNTP en de fluorescente label is verbroken en de label wordt vrijgesteld. Stap 4: De polymerase verplaatst zich naar de volgende positie. Stap 5: Het proces wordt herhaald (Korlach et al., 2010).
Het concept van Single-Molecule, Real-Time (SMRTTM) DNA-sequencing geïllustreerd op Figuur 7, steunt daarnaast ook op het principe van zero-mode waveguides (ZMW). ZMW-arrays bevatten gaten van gemiddeld 100nm diameter die zeer dicht opeen geplaatst zijn. Een enkele ZMW fungeert als een nanofotonische visualisatiekamer waarin de polymerisatiereactie uitgevoerd door een enkele polymerase molecule kan opgevolgd worden. De inhoud van een ZMW-kamer is gemiddeld 100 zeptoliter (10-21L). Het klein volume maakt het onderscheiden
van een fluorescent signaal door diffusie en een signaal door het inbouwen van een dNTP door de polymerase mogelijk (Korlach et al., 2010).
Het voornaamste verschil en voordeel van PacBio ten opzichte van Illumina sequencing is de mogelijkheid om een groot aantal reads van aanzienlijke grootte (tot 3kb) te verkrijgen. Dankzij de langere reads kunnen zogenaamde gaten in het genoom, delen waarvan de sequentie nog niet werd bepaald, vervolledigd worden. Daarnaast kan de lengte van regio’s met meerdere repeats van een bepaalde sequentie beter gevisualiseerd worden.
1.2.2 Viruseliminatiemethoden
Een veel gebruikte strategie voor het bekomen van virusvrije gewassen is thermotherapie gecombineerd met meristeemculturen. Meristemen worden hierbij verkozen omdat de floëem- en xyleemvaten van een plant niet tot daar reiken (Al-Taleb et al., 2011). Dit maakt van de meristemen het deel van de plant waar de concentratie aan virussen minimaal is. Daarnaast werd in vorige studies reeds aangetoond dat de virusconcentratie in een plant niet gelijk verdeeld is en daalt naar de uiteinden toe (zowel top- als wortel-apex) (Mori, 1971).
Thermotherapie bestaat uit het opgroeien van scheuten tot kleine plantjes uit knollen in geval van knolgewassen of uit stekken van planten bij een hogere temperatuur en vochtigheidsgraad. De verhoogde temperatuur wordt gebruikt om de synthese van viruspartikels te beletten. In principe worden viruspartikels continu geproduceerd en gedegradeerd. Bij een verhoogde temperatuur, tussen 30-40°C, wordt de synthese stopgezet maar gaat de degradatie alsnog door (Walkey, 1991). De virussen worden echter niet volledig geïnactiveerd. De temperatuur die hiervoor nodig zou zijn varieert van 40-90°C, wat niet realistisch is voor een plant. Door jonge geïnfecteerde planten te laten opgroeien bij een verhoogde temperatuur, zullen de nieuwe delen van de plant geen viruspartikels meer bevatten, maar de onderliggende delen wel. De meristemen moeten dan in vitro geïnitieerd worden om verder tot een plant uit te groeien. De combinatie van thermotherapie met meristeemculturen werd reeds beschreven in talrijke studies, waarvan enkele hieronder beschreven.
Vivek & Modgil (2018) hebben een artikel gedupliceerd waarin beschreven wordt hoe virusvrije appel cultivars bekomen werden door middel van thermotherapie gevolgd door meristeemculturen. Hiervoor werden geïnfecteerde scheuten verzameld op verschillende appelbomen die vervolgens gedurende vier weken bij 37-40°C werden geïncubeerd. Na thermotherapie werden de apicale en laterale knoppen van de scheuten verwijderd en gesteriliseerd. Vervolgens werden de meristemen uit de knoppen gesneden, waarbij 1-4 bladprimordia werden behouden. Na 6 weken werden de overlevende culturen uitgezet op nieuw medium. Bladstalen werden genomen voor DAS ELISA analyse om na te gaan of de oorspronkelijke virussen (apple chlorotic leaf spot virus en apple stem pitting virus) succesvol verwijderd werden. Het resultaat van de studie concludeerde dat bovengenoemde virussen geëlimineerd konden worden dankzij de combinatie van thermotherapie en meristeemculturen. De grootte van de meristemen heeft echter een invloed op de concentratie aan virussen teruggevonden in de nieuwe scheuten. Hoe groter het meristeem, hoe groter de overlevingskans ervan maar hoe kleiner de kans dat de aanwezige virussen succesvol werden verwijderd. Deze technieken werden ook reeds toegepast op yam, namelijk D. alata, voor de eliminatie van
Flexous Rod virussen (Mantell et al., 1980). Hierbij werden stukken van yamknollen geplaatst
in een hittekamer bij 36 ± 1°C en 90 ± 5% relatieve vochtigheid gedurende twee tot drie weken. Vervolgens werden apicale wortel- en scheutmeristemen uitgesneden van 16-28 weken oude planten, opgegroeid uit de knollen behandeld door thermotherapie. Slechts 19% van de meristemen (112) konden uitgroeien tot scheuten. De doeltreffendheid van de behandeling werd nagegaan door symptoom en elektronenmicroscoop observaties. De afwezigheid van deeltjes van het Flexous Rod virus en cytoplasmatische inclusies tonen aan dat het toepassen van thermotherapie in combinatie met meristeemculturen effectief doeltreffend is voor het uitroeien van potyvirus infecties in D. alata.
Filloux et al. (2006) hebben eveneens Dioscorea spp. getarget voor viruseliminatie waarbij de focus lag op badna- en potexvirussen, het yam mild mosaic virus (YMMV) en het yam mosaic virus (YMV). Geïnfecteerde in vitro yam plantjes werden geplaatst in een incubator bij 34-36°C overdag en 32°C ’s nachts gedurende 45-60 dagen tot de in vitro plantjes 2 tot 6cm groot waren. De fotoperiode was 12u/dag. Na thermotherapie werden 20 apicale meristemen weggesneden onder een binoculaire microscoop. De meristemen werden gedurende twee maanden opgegroeid waarna ze opnieuw werden getest op de aanwezigheid van virussen. In de geregenereerde planten konden bij 2% van de gevallen de badnavirussen worden verwijderd, 60% van de Potexvirussen, 46% van de YMMV en 21% van de YMV. Al zorgt thermotherapie in combinatie met meristeemculturen niet voor de volledige verwijdering van alle aanwezige virussen is het toch mogelijk om enkele virusvrije culturen te bekomen. Door vermeerdering kunnen nadien talrijke virusvrije culturen worden bekomen.