GEÏNDIVIDUALISEERD TRAINEN: ROL
SPIERVEZELVERDELING IN HET
BEPALEN VAN TRAININGSMODALITEITEN
Aantal woorden: 20.188Sam De Cooman, Chloé Decooman
Studentennummer: 01500012, 01504899
Promotor: Prof. Dr. Wim Derave Copromotor: Dr. Laura Blancquaert
A. VOORWOORD
Zo’n twee jaar geleden kregen wij het onderwerp van onze masterproef te horen. Dit was een heugelijk moment aangezien dit onderwerp bovenaan ons lijstje stond, omwille van de grote relevantie binnen de sportpraktijk. Het gaat om nieuwe, en daarom net zo uitdagende en aantrekkelijke, materie binnen de fysiologie.
Een voorwoord zonder bedanking, is zoals het HILO zonder Rudi, namelijk niet compleet. Daarom gaat onze grootste dank uit naar onze begeleidster drs. Laura Blancquaert, die ons steeds heeft voorzien van opbouwende, doch kritische feedback die onze masterproef tot een hoger niveau heeft getild. Verder bedanken we ook graag prof. Derave. Deze staat niet alleen bekend als een gevestigde waarde binnen de (sport)fysiologie, maar dezer dagen ook als ‘de bezorgde professor’. Trots konden wij tegen onze naasten zeggen, ‘hé kijk, dat is onze
promotor’.
Dit brengt ons tot de uitbraak van de COVID-19 pandemie in maart 2020. Alhoewel dit gebeuren roet in het eten gooide voor sommige analyses, heeft dit onze samenwerking niet teniet gedaan. Een tiental videogesprekken per dag, zonder daarbij te overdrijven, heeft de samenwerking steeds vlot laten verlopen.
Vervolgens een warm dankjewel aan de totale 18 proefpersonen die hebben deelgenomen aan onze studie. Een twintigtal keren zijn jullie afgezakt naar ons sportlabo hier te Gent. Zelfs meerdere keren afzien op de fiets en het afnemen van de spierbiopten hield jullie niet tegen (alhoewel Chloé daar iets minder goed tegen kon om 8 uur ‘s ochtends). Eveneens ‘nen dikke
merci’ aan Maïté Leleu voor het nalezen van onze discussie.
Tenslotte hadden wij elkaar nog graag bedankt. “Bedankt Chloé voor jouw creativiteit. Bedankt om mij af en toe enige realiteitszin bij te brengen en mijn soms (te) vergaande ideeën in te perken. Maar bovenal bedankt voor jouw inzet de laatste maanden. Dedie van Giesbaargen
emmen da goe gedoan!” (Sam). “Bedankt Sam voor mijn thesispartner te zijn. Je hebt mij steeds
in de goede richting geduwd, ook wanneer ik het even moeilijk had. Jouw denkvermogen en inzicht in bepaalde zaken heeft ook mij intellectueel geprikkeld en geholpen in deze scriptie. Verder zou ik graag nog mijn beide huisgenoten bedanken die al mijn ups en downs van dichtbij hebben meegemaakt en hierbij een echte rots in de branding waren. Ten slotte, de grootste steun en toeverlaat binnen deze opleiding: bedankt Lexie, Delfje, Florinski en Feline voor de eindeloze videogesprekken en ontspanningsmomenten.” (Chloé).
B. ABSTRACT
Inleiding: Anno 2020 wordt in zowel individuele sporten als teamsporten gestreefd naar een doorgedreven individualisatie van trainingsmodaliteiten om prestatie te optimaliseren. Een onderscheid maken in trainingsmodaliteiten tussen atleten op basis van hun spiervezeltypologie is tot op heden een ongekende vorm van individualisering. Het doel van deze thesis bestond er dan ook uit om na te gaan of een zes weken durende hoge intensiteit interval trainingsperiode (HIIT) een verschillende respons impliceert tussen twee groepen met een tegenovergestelde spiervezeltypologie. Aangezien snelle spiervezels verondersteld worden meer adequaat gestimuleerd te worden bij hogere intensiteiten vergeleken met trage spiervezels, werd gehypothetiseerd dat de snelle groep een grotere verbetering zouden bewerkstelligen in VO₂max en andere cardiorespiratoire parameters.
Methode: Aan de hand van de ‘Muscle Talent Scan’ werden 9 mannelijke, sedentaire proefpersonen opgedeeld in een trage en een snelle groep. Beide groepen werden aan eenzelfde zes weken durende HIIT-interventie op een fietsergometer onderworpen. Hierbij werden de intensieve intervallen afgewerkt aan 100% van het respiratoir compensatiepunt. Deze werden afgewisseld met een actieve recuperatie aan 90% van de gasuitwisseling drempel. Het volume werd overheen de weken verhoogd. Een maximale inspanningstest met een 25 W/min rampprotocol werd uitgevoerd voorafgaand, halfweg en na de HIIT-interventie. Een submaximale inspanningstest werd voor en na de trainingsperiode uitgevoerd.
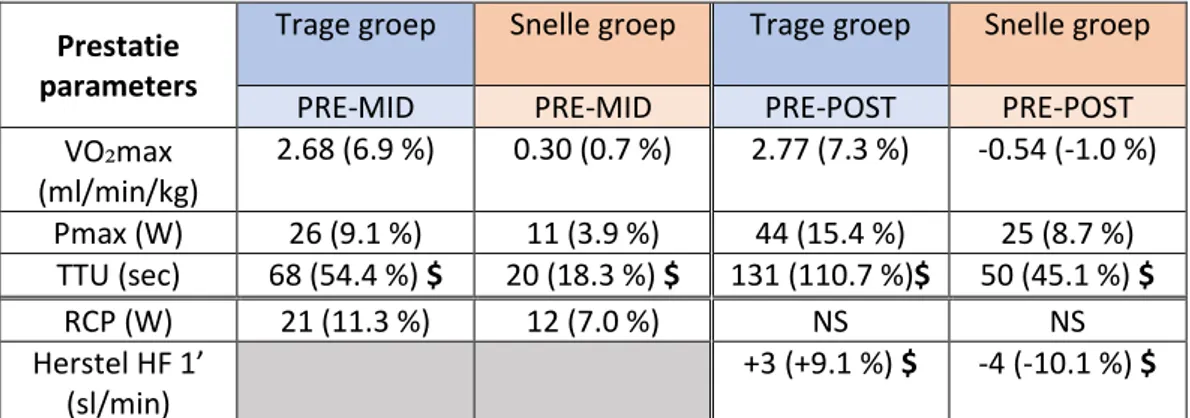
Resultaten: De trage groep kon een significant grotere stijging in VO₂max bewerkstelligen ten opzichte van de snelle groep (p = 0.009). Hierbij slaagde de eerstgenoemde erin een toename van 7.3 % (p = 0.003) te realiseren, terwijl de laatstgenoemde met -1.0 % geen significante verandering (p = 0.872) kon verwezenlijken na het doorlopen van de trainingsperiode. Daarentegen lag voor beide groepen het maximaal vermogen uit de maximale inspanningstest na de interventie significant hoger ten opzichte van de test bij aanvang van de trainingsperiode (trage groep: 15.4 %,; snelle groep: 8.7 %, p < 0.05). Echter was ook voor deze parameter de toename bij de trage groep groter (p = 0.003). Voor submaximale parameters bestond op groepsniveau een minder uitgesproken verschil tussen de trage en snelle groep. Enkel voor herstel in hartslag één minuut na de submaximale test werd een trend tot significant verschil bevonden tussen de groepen (p = 0.094). Binnen de snelle groep lijkt een groter aandeel proefpersonen non-respons te vertonen op verscheidene prestatieparameters in vergelijking met de trage groep. Wel neemt het aantal proefpersonen dat non-respons vertoont binnen de trage groep toe overheen de laatste drie weken van de trainingsperiode.
Conclusie: De voornaamste bevinding van dit onderzoek was dat de spiervezeltypologie van individuen aan de basis kan liggen voor een verschil in respons op training. Desondanks uit voorgaande literatuur reeds bleek dat snelle spiervezels meer adequaat gestimuleerd worden bij hogere intensiteiten, waren er meer uitgesproken effecten in de trage groep ten opzichte van de snelle groep voor VO₂max en verschillende andere prestatieparameters. De bevindingen uit deze studie tonen aan dat een mogelijks nieuwe factor is geïdentificeerd om variatie in trainingsrespons te verklaren, namelijk spiervezeltypologie.
C. INHOUDSTABEL
A. VOORWOORD ... I B. ABSTRACT ... III C. INHOUDSTABEL ... V D. AFKORTINGENLIJST ... VII I. LITERATUUR ... 3 1. Situering……..……….3 2. Spiervezels………...4 2.1. Spiercontractie ... 4 2.2. Spiervezeltypes ... 42.3. Determinanten spiervezeltypesamenstelling: nature vs nurture... 6
2.4. Kwantificatie spiervezels ... 10
3. Training………..………...………….11
3.1. Trainingsmodaliteiten ... 11
3.2. Trainingsvormen en rekrutering spiervezels ... 12
3.3. Aanpassingen aan training ... 13
4. Variatie in respons op training………...…20
4.1. Probleem van het groepsgemiddelde ... 20
4.2. Intra-individuele variatie respons op training bij verschillende trainingsvormen..19
4.3. Non-responders ... 22
4.4. Oorzaken variatie in respons op training ... 23
4.5. Bestaande individualisering ... 26 5. Trainingsprogramma en spiervezeltype……….27 5.1. Vermoeidheid en herstel ... 27 II. Methode ... 35 1. Populatie……….…………35 2. Experimentele procedure……….…...……36 2.1. Protocol ¹H-MRS ... 36 2.2. Studiedesign ... 38 2.3. Spierbiopt ... 38 2.4. Maximale inspanningstest ... 39 2.5. Submaximale inspanningstest ... 40 2.6. Trainingsperiode ... 40 3. Meetinstrumenten……….………..…41
3.1. Antropometrie ... 41 3.2. Gasuitwisseling en hartfrequentie ... 41 3.3. Fietsergometrie ... 42 3.4. Bloedlactaat ... 42 4. Data-analyse……….………..42 4.1. Maximale inspanningstest ... 42 4.2. Trainingsbelasting ... 45
4.3. Groot mechanische efficiëntie en substraatverbruik ... 45
4.4. Spiervezeldistributie ... 46
III. RESULTATEN ... 51
1. Pre-training karakteristieken………..51
2. Effect training op cardiorespiratoire parameters uit maximale inspanningstest………52
2.1. VO₂max ... 52
2.2. Pmax ... 54
2.3. Inspanningsdrempels ... 55
2.4. Tijd tot uitputting ... 56
3. Effect training op cardiorespiratoire parameters uit submaximale inspanningstest…...57
3.1. HF submax ... 57
3.2. Lactaat ... 58
3.3. RPE en %VO₂Max ... 58
3.4. Substraatverbruik en GME ... 59
IV. DISCUSSIE ... 65
1. Invloed van SVT op respons training………65
2. Vergelijking in toename van de parameters t.o.v. andere trainingsinterventies………71
3. Individuele respons………74
4. Limitaties………...79
V. CONCLUSIE ... 83
VI. BIBLIOGRAFIE ... 84
D. AFKORTINGENLIJST
ADP: adenosinedifosfaat ATPase: adenosine trifosfatase bTRIMP: Banister Training Impulse DT: duurtraining
EDT: extensieve duurtraining eTRIMP: Edwards Training Impulse EIT: extensieve intervaltraining GET: gasuitwisseling drempel GME: groot mechanische efficiëntie HF: hartfrequentie
HIIT: hoge intensiteit intervaltraining
H-MRS: Proton Magnetische Resonantie Spectroscopie H-MRS: proton magnetische resonantie spectroscopie HRV: hartslagvariabiliteit
IDT: intensieve duurtraining
iEMG: geïntegreerde elektromyografie IIT: intensieve intervaltraining
IT: inspanningstest IT: intervaltraining LG: lichaamsgewicht LL: lichaamslengte LT: lactaat drempel
luTRIMP: Lucia Training Impulse ME: motor eenheid/motor eenheden MHC: myosine zware keten
MRS: magnetische resonantie spectroscopie MRT: mean response time
PETCO₂: de fractie koolstofdioxide in de uitgeademde lucht PETO₂: de fractie zuurstof in de uitgeademde lucht
Pi; fosfaat-ion
Pmax: maximaal vermogen RCP: respiratoir compensatiepunt SD: standaard deviatie
SDH: succinaat-dehydrogenase SIT: sprint intervaltraining Sl/min: slagen per minuut
sRPE: session ratings of perceived exertions ST: sprinttraining
SV: spiervezel/spiervezels
SVT: spiervezeltype/spiervezeltypologie TEM: typical error of measurements TL: trainingsbelasting
TTU: tijd tot uitputting
VCO₂: uitgeademde koolstofdioxide VE/VCO₂: equivalent van VCO₂ respons VE/VO₂: equivalent van VO₂ respons VE: ventilatie
VL: vezellengte
Vo: maximale verkortingssnelheid VO₂: zuurstofopname
VO₂max: maximale zuurstofopname α MN: α-motorneuron
I.
LITERATUUR
1. Situering
Individualisering is een ‘hot topic’ in de topsportwereld. Iedere duursporter die zichzelf serieus neemt traint aan de hand van een op maat gemaakt trainingsprogramma. In zo een trainingsprogramma wordt op basis van de doelstellingen van een atleet bepaalde trainingsvormen met specifieke modaliteiten voorgeschreven. Een wielrenner die wil uitblinken in de voorjaarsklassiekers zal van zijn fysieke trainer voornamelijk duurtrainingen voorgeschoteld krijgen. Een loper die de 100m spurt op een provinciaal kampioenschap wil winnen, zal daarentegen vaker kortere inspanningen aan hoge intensiteiten uitvoeren in zijn trainingen. Op basis van vooropgestelde doelen wordt dus met andere woorden vaak gegrepen naar specifieke trainingsvormen om de doelstelling te behalen.
Uit de literatuur blijkt echter dat wanneer een groep subjecten een trainingsprogramma met een bepaalde trainingsvorm volgt, er een grote variatie in trainingseffect kan plaatsvinden tussen subjecten. Dit betekent dus dat bij twee wielrenners die samen trainen voor de voorjaarklassiekers en zeer veel duurtrainingen doen aan lage intensiteit, het wel eens zou kunnen zijn dat de ene een veel groter trainingseffect heeft dan de andere, desondanks ze dezelfde trainingen uitvoerden. Erfelijkheid speelt een belangrijke rol in deze variatie in respons op training. Het valt dan ook op dat een aanzienlijk aantal topsporters een voorganger hebben binnen de familie die ook uitblonk in dezelfde discipline. De rol van spiervezeltypering van een atleet inzake het excelleren in bepaalde disciplines werd reeds vaak besproken en is van significant belang. De rol van deze spiervezeltypering in de variatie in respons op training is echter een vrijwel onbesproken onderwerp in de literatuur. Het doel van deze studie bestaat er dan ook uit om na te gaan of twee groepen met een tegenovergestelde spiervezeltypologie een verschillende respons vertonen in VO₂max en andere cardiorespiratoire parameters na een trainingsinterventie.
2. Spiervezels
2.1. Spiercontractie
Een spiervezel (SV) is opgebouwd uit myofibrillen bestaande uit bundels dikke en dunne myofilamenten. De hoofdbestanddelen van deze myofilamenten zijn actine en myosine waarbij actine wordt teruggevonden in de dunne filamenten, terwijl myosine de bouwsteen is van de dikke filamenten. Een spiercontractie vindt plaats door het actief verkorten van deze myofibrillen, wat men het mechanisme van de glijdende filamenten noemt. Elke actinemolecule heeft een actieve plaats die in staat is om te binden met de myosinekoppen in aanwezigheid van calcium. Bij deze binding wordt energie vrijgesteld door de hydrolyse van ATP waardoor er een knik ontstaan in de myosinekoppen met een verkorting van de myofibrillen als gevolg. De trigger voor deze spiercontractie is een excitatie van een α-motorneuron (α MN) door een signaal vanuit de hersenen. Elke SV wordt geïnnerveerd door een tak van een α MN waarbij het totaal van één α-motorneuron en zijn bijhorende SV een motorische eenheid (ME) wordt genoemd. Één enkel α MN stuurt meerdere SV aan waarbij een onderscheid wordt gemaakt tussen een kleine ME, bestaande uit enkele SV, en een grote ME, gaande tot wel 2000 SV per ME (Feinstein et al., 1955). De actiepotentiaal bepaalt of er een kleine of grote ME wordt aangesproken. Kleinere ME hebben een kleinere drempelwaarde in vergelijking met grote ME.
2.2. Spiervezeltypes
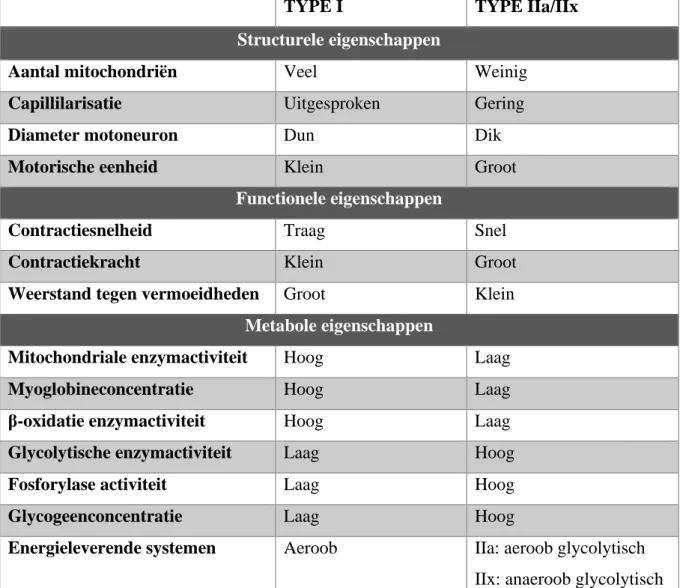
De SV worden al sinds enkele decennia opgesplitst in twee soorten op basis van eigenschappen van de myosine zware keten (MHC), welk een deel is van een eiwit. De eerste soort is geschikt voor het leveren van duurinspanningen en worden de trage SV genoemd of ook wel de type I SV. Dit zijn traag samentrekkende, oxidatieve SV en hebben dus een aerobe energielevering. Een tweede soort, type II SV, zijn dan weer meer geschikt voor kortstondige, maximale inspanningen. Deze bereiken na prikkeling hun maximale contractiekracht na 0.01 seconden, wat drie keer zo snel is als type I vezels. Type II vezels worden dan ook logischerwijs gelabeld als de snelle SV (Martini &
SV-type ook gerekruteerd worden bij aerobe inspanningen. Type IIx is snel en glycolytisch. Dit type is bijgevolg vooral geschikt voor zeer korte, maximale inspanningen. Hybride vezels zijn een laatste soort SV welke een co-expressie van minstens twee of drie soorten MHC-isovormen vertonen. Dit type SV wordt gesuggereerd SV in de overgangsfase te zijn.
Tabel 1: eigenschappen van SV (gebaseerd op Bourgois, 2017-2018)
TYPE I TYPE IIa/IIx
Structurele eigenschappen
Aantal mitochondriën Veel Weinig
Capillilarisatie Uitgesproken Gering
Diameter motoneuron Dun Dik
Motorische eenheid Klein Groot
Functionele eigenschappen
Contractiesnelheid Traag Snel
Contractiekracht Klein Groot
Weerstand tegen vermoeidheden Groot Klein
Metabole eigenschappen
Mitochondriale enzymactiviteit Hoog Laag
Myoglobineconcentratie Hoog Laag
β-oxidatie enzymactiviteit Hoog Laag
Glycolytische enzymactiviteit Laag Hoog
Fosforylase activiteit Laag Hoog
Glycogeenconcentratie Laag Hoog
Energieleverende systemen Aeroob IIa: aeroob glycolytisch
2.3. Determinanten spiervezeltypesamenstelling: nature vs nurture
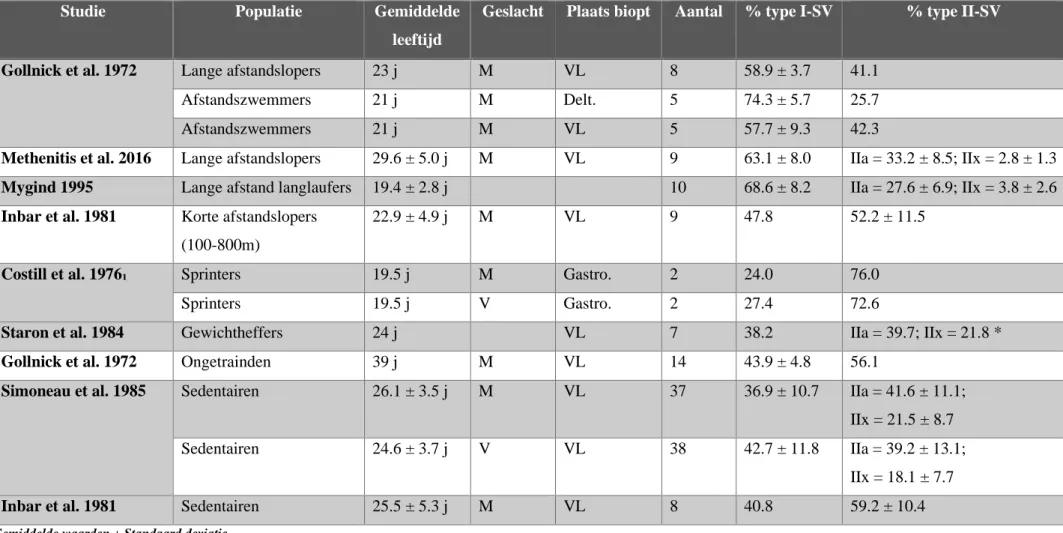
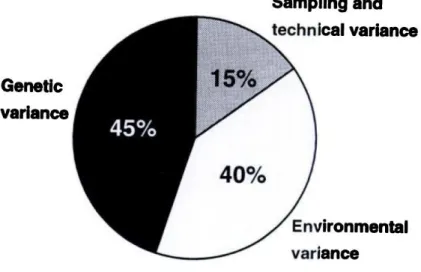
Overheen de algemene populatie is er een grote variatie inzake spiervezelcompositie. Naast genetische aanleg (45%) zijn ook het niveau van fysieke activiteit, voedingsgewoonten en levensstijl (omgevingsfactoren (40%)) oorzaken hiervoor (zie figuur 1, Simoneau & Bouchard, 1995). Getrainde uithoudingsatleten worden getypeerd door een groter percentage trage SV en dus ook een beter aeroob vermogen vergeleken met sprinters, werpers, ver- en hoogspringers (zie tabel 2). Het feit dat een atleet in een bepaalde sportdiscipline excelleert komt voort uit een vorm van natuurlijke selectie en kan worden teruggekoppeld naar de spiervezeltypologie van de atleet. Dat spierdistributie zeer uiteenlopend is in de globale populatie, blijkt uit de resultaten van Staron et al. (2000). Deze onderzoekers verzamelden gegevens van 150 jonge mannen en vrouwen en toonden aan dat het bereik van SV type I reikt van 16.5 tot 97.4% bij vrouwen en van 17.6% tot 65.6% bij mannen. Wanneer we intra-individueel kijken kan er een intermusculaire relatie gevonden worden in SV compositie. Dit betekent dat bij personen die in een bepaalde spier een relatief hoog percentage type I vezels vertonen, er ook in andere spieren een tendens zal zijn naar een dominante aanwezigheid van type I vezels (Vikne et al., 2012).
Tabel 2: spiervezeldistributie bij verschillende populaties
Studie Populatie Gemiddelde
leeftijd
Geslacht Plaats biopt Aantal % type I-SV % type II-SV
Gollnick et al. 1972 Lange afstandslopers 23 j M VL 8 58.9 ± 3.7 41.1 Afstandszwemmers 21 j M Delt. 5 74.3 ± 5.7 25.7 Afstandszwemmers 21 j M VL 5 57.7 ± 9.3 42.3
Methenitis et al. 2016 Lange afstandslopers 29.6 ± 5.0 j M VL 9 63.1 ± 8.0 IIa = 33.2 ± 8.5; IIx = 2.8 ± 1.3
Mygind 1995 Lange afstand langlaufers 19.4 ± 2.8 j 10 68.6 ± 8.2 IIa = 27.6 ± 6.9; IIx = 3.8 ± 2.6
Inbar et al. 1981 Korte afstandslopers (100-800m)
22.9 ± 4.9 j M VL 9 47.8 52.2 ± 11.5
Costill et al. 1976₁ Sprinters 19.5 j M Gastro. 2 24.0 76.0
Sprinters 19.5 j V Gastro. 2 27.4 72.6
Staron et al. 1984 Gewichtheffers 24 j VL 7 38.2 IIa = 39.7; IIx = 21.8 *
Gollnick et al. 1972 Ongetrainden 39 j M VL 14 43.9 ± 4.8 56.1
Simoneau et al. 1985 Sedentairen 26.1 ± 3.5 j M VL 37 36.9 ± 10.7 IIa = 41.6 ± 11.1; IIx = 21.5 ± 8.7 Sedentairen 24.6 ± 3.7 j V VL 38 42.7 ± 11.8 IIa = 39.2 ± 13.1;
IIx = 18.1 ± 7.7
Inbar et al. 1981 Sedentairen 25.5 ± 5.3 j M VL 8 40.8 59.2 ± 10.4
Gemiddelde waarden ± Standaard deviatie.
M = mannen; V = vrouwen; j = jaar; VL = m. vastus lateralis; Delt. = m. deltoideus; Gastro. = m. gastrocnemius *(IIc niet mee opgenomen)
2.3.1. Leeftijd
Ouder worden induceert heel wat veranderingen aan het lichaam. Ook in de skeletspier vinden zowel morfologische als functionele aanpassingen plaats (Porter et al., 1995). Veroudering zorgt voor een daling in het aandeel type II SV (Larsson et al., 1978). Daarnaast wordt er een toename in het aantal hybridevezels geobserveerd met als vaakst voorkomende de co-expressie van MHC I en IIa (Andersen et al., 1999). Een mogelijke verklaring voor de stijging in hybridevezels bij ouderen is de reïnnervatiehypothese. Deze houdt in dat bij ouderen een verlies van motoreenheden leidt tot denervatie, gevolgd door reïnnervatie. Door deze reïnnervatie ontvangt een meervoudig aantal vezels in de vezelpool een andere neurale input die verschillend is van hun myogene afkomst. Dit zorgt voor een tegenstrijdig signaal voor de vezel en geeft uiteindelijk aanleiding tot gemengde expressie van MHC-isovormen in de SV (Andersen et al., 1999).
2.3.2. Geslacht
Het is moeilijk om een verschil in spiervezelcompositie te beschrijven op basis van geslacht, aangezien in de literatuur hieromtrent contesterende resultaten te vinden zijn. Meerdere studies tonen dat op basis van geslacht geen verschil gevonden wordt in spiervezelcompositie (Toft et al., 2003; Sale et al., 1987₂; Coggan et al., 1992). Wanneer er echter toch van een verschil gesproken wordt, vertonen vrouwen een groter percentage type I SV ten opzichte van mannen (Simoneau & Bouchard, 1989; Haizlip et al., 2015). Verschillen in leeftijd, profiel van de proefpersoon en methodologie kunnen aan de basis liggen voor de uiteenlopende bevindingen (Staron et al., 2000).
2.3.3. Genen en etniciteit
Simoneau & Bouchard (1995) observeerden dat ongeveer 25% van de Noord-Amerikaanse bevolking ofwel minder dan 35% ofwel meer dan 65% type I SV bevat. Spierbiopten van 32 paar broers, 26 paar mannelijke en vrouwelijke twee-eiige tweelingen en 35 paar mannelijke en vrouwelijke eeneiige tweelingen werden vergeleken. Hierbij kon 45% van de variantie in proportie van type I SV toegeschreven worden aan genotype (zie figuur 1, uit Simoneau & Bouchard, 1995).
zwart Afrikaanse groep, welke een groter percentage type IIa vezels bezitten (6.7%,p<0.05). Er werd geen verschil opgemerkt tussen beide groepen voor type IIx (Ama et al., 1986).
Figuur 1: verklaring variatie in proportie type I SV (Uit SImoneau & Bouchard, 1995)
2.3.4. Shift door training
Een vaak bediscussieerd onderwerp in de literatuur is de invloed van training op de spiervezelcompositie. Simoneau & Bouchard (1995) toonden aan dat naast genetische aanleg ook omgeving voor 40% meespeelt in variatie van SV distributie (zie figuur 1, uit Simoneau & Bouchard, 1995). Studies van Gollnick et al. (1973₁), Bagby et al. (1972); Saltin et al. (1976) zagen geen verschil in SV-compositie na een trainingsperiode, waar verschillende trainingsvormen werden opgelegd. Dit impliceert dat de genetische factor een grote rol speelt. Hierbij moet wel vermeld worden dat Gollnick et al. (1973₁) geen onderscheid maakte tussen de verschillende type II vormen en dus enkel rekening hield met de shift van type II naar type I. Desalniettemin zijn er studies die het tegendeel aantonen en wel een shift door training vaststellen. Een significante afname (p<0.05) van IIx en een toename in IIa geïnduceerd door duurtraining werd waargenomen in de studies van Green et al. (1979); Andersen & Henriksson (1977); Coggan et al. (1992) en Harber et al. (2012). Studies die een shift van type I naar type II beschrijven bestaan, maar zijn eerder gering (Andersen et al., 1994; Jansson et al. 1990). De shift van type II naar type I werd reeds aangetoond via elektrische stimulatie van de spier (Jolesz & Sreter, 1981; Salmons & Henriksson 1981) maar de literatuur is inconsistent over het bestaan van dergelijke shift in spiervezelcompositie geïnduceerd door training.
2.4. Kwantificatie spiervezels
2.4.1. Spierbiopten
Spierbiopten zijn al enkele decennia de gouden standaard wanneer het gaat over de bepaling van spiervezeltype. Een spierbiopt wordt genomen met behulp van een holle naald. Via deze methode wordt onder plaatselijke verdoving een stukje spierweefsel gehaald ongeveer halverwege tussen de oorsprong en de insertie van de spier (Lexell et al., 1985; Bergström, 1975). De vastus lateralis is de voorkeursspier voor biopten vanwege de gemengde spiervezelsamenstelling, de trainbaarheid en de toegankelijkheid (Bergström, 1975),. Het is een invasieve methode, wat resulteert in schade in de desbetreffende spier. Een biopt omvat 10-200 mg spier, wat overeenkomt met slechts 0.01% van de totale spiermassa (Albracht et al., 2008) en bevat slechts enkele honderden vezels (Baguet et al., 2011). Lexell et al. (1985) suggereren daarom dat een spierbiopt niet noodzakelijk representatief is voor de gehele spier. De distributie van de soorten spiervezeltypes veranderen naarmate de diepte van de spier. Deze studie vond dat de proportie type I vezels stijgt, wanneer diepere spiervezels worden vergeleken met meer oppervlakkige vezels.
De indeling van de SV in de verschillende categorieën van spiervezeltypes kan op een aantal manieren gebeuren. Een histochemische kleuring van myofibrillaire adenosine trifosfatase (ATPase) op cryostaatsecties van bevroren weefsel (Padykula & Herman 1955; Brooke & Kaiser 1970) is de traditionele methode. Hierbij resulteert de chemische reactie van ATPase op myosine met cobalt in een zwarte kleur. Naarmate er een hoge of lage pH aanwezig is verandert de kleur. Bij een zure incubatie vertoont type I een zwarte kleur, bij een basische pH vertoont vooral type II een zwarte, meer donkere kleur. Als deze methode wordt gebruikt spreekt men voornamelijk over type IIb i.p.v. type IIx, omdat dit al een oudere techniek is. Andere populaire methoden zijn bijvoorbeeld de immunohistochemische techniek waarbij specifieke antilichamen gaan binden met MHC-isovormen (MHC-I, MHC-IIa en MHC-IIx), of via gel elektroforese, waarbij de verschillende MHC-isovormen onderscheiden kunnen worden op basis van moleculair gewicht (Scott et al., 2001)
carnosine op een indirecte manier de spiervezeltypesamenstelling in te schatten. Er bestaat een sterke positieve correlatie (r=0.714, p<0.01) tussen carnosine concentratie en de relatieve oppervlakte type II vezels in de gastrocnemius (Baguet et al., 2011). Aan de hand van een database inzake de carnosine-inhoud van 83 niet specifiek getrainde subjecten is het mogelijk om ieders carnosine concentratie op basis van Z-scores hiermee te vergelijken en zo een schatting van de spiervezeldistributie te maken. In contrast met de klassieke spierbiopt-methode wordt bij deze techniek geen schade veroorzaakt aan de spier. Daarnaast is deze methode ook minder arbeids-en tijdsintensief en wordt een meer globaal beeld verschaft over de spiervezelcompositie van de spier. Dit in tegenstelling tot de 0.01% van de totale spier die onderzocht wordt op basis van enkele honderden vezels bij een klassieke spierbiopt (Baguet et al., 2011; Albracht et al., 2008). Een nadeel van de ¹H-MRS is dat het gebaseerd is op de kwantificatie van slecht 1 metaboliet, namelijk carnosine. Chronisch vegetarisme (Everaert et al., 2011), alsook beta-alanine supplementatie (Derave et al., 2007) kunnen de spiercarnosine concentratie beïnvloeden, met een foutieve bepaling van de spiervezelcompositie tot gevolg.
3. Training
3.1. Trainingsmodaliteiten
Heden ten dage bestaan tal van trainingsvormen. Hieronder kunnen we duurtraining (DT), intervaltraining (IT), sprinttraining (ST) en een combinatie van voorgaande onderscheiden. Elk trainingsprogramma heeft zijn eigen specifieke trainingsmodaliteiten. Deze belastingscomponenten worden kwantitatief vastgelegd en gedefinieerd als volgt: de trainingsintensiteit is de sterkte van de trainingsprikkel, ofwel de geleverde arbeid per tijdseenheid. Dit kan gezien worden als het percentage van de maximale hartslag/snelheid/kracht. De trainingsfrequentie is het aantal trainingseenheden per tijdseenheid. Het trainingsvolume is de totale hoeveelheid werk die per trainingssessie wordt verricht. De combinatie van deze modaliteiten zal de uiteindelijke trainingsvorm bepalen. Algemeen kan worden gesteld dat er een inverse relatie bestaat tussen de intensiteit en het volume van een training.
3.2. Trainingsvormen en rekrutering spiervezels
3.2.1. Size principle
Kleine ME worden zoals reeds vermeld getypeerd door een lagere drempelwaarde in vergelijking met grotere ME. Wanneer een bepaalde taak slechts een beperkte mate van spierkracht vereist, zijn het enkel deze kleine ME die gerekruteerd worden. Wanneer de intensiteit van de taak wordt verhoogd en een grotere krachtlevering nodig is, zal dit de rekrutering van grotere ME (en dus snelle SV) met zich meebrengen (Haff et al., 2001) (zie grafiek 1, naar Willmore & Costill, 1994). Naast toenemende intensiteit is ook de snelheid waarmee de spier contraheert positief gecorreleerd met de rekrutering van grotere ME (Sale, 1987₁). Ook vermoeidheid lijkt een rol te spelen inzake de spiervezelrekrutering. Waar bij een inspanning aan een lage intensiteit aanvankelijk uitsluitend beroep wordt gedaan op kleine ME, zal naarmate de inspanning langer duurt vermoeidheid van type I vezels optreden. Dit heeft als gevolg de rekrutering van type II vezels. (Gollnick et al., 1973₂). Zeer explosieve taken zoals bijvoorbeeld kogelstoten, waarbij een zeer grote kracht zo snel mogelijk gegenereerd moet worden, lijken dit rekruteringspatroon niet te volgen. Hierbij contraheren grotere ME voor de kleine ME (Desmedt & Godaux, 1977).
3.2.2. Duurtraining
DT bestaat uit langdurige, ononderbroken inspanningen. DT kan men verder opsplitsen in extensieve duurtraining (EDT) en intensieve duurtraining (IDT). Bij EDT gaat het om een groot volume met een lage intensiteit die zich situeert rond de aerobe drempel. Tijdens EDT worden voornamelijk type I vezels gerekruteerd (Gollnick et al., 1974). De intensiteit bij IDT ligt hoger en situeert zich eerder tussen de aerobe en anaerobe drempel. Deze trainingsvorm rekruteert bijgevolg meer type IIa vezels in vergelijking met EDT (Costill et al., 1973).
3.2.3. Intervaltraining
IT wordt gekenmerkt door de afwisseling van inspanningen met hoge en lage intensiteit. Men onderscheidt extensieve intervaltraining (EIT) en intensieve intervaltraining (IIT). Onder deze laatst genoemde kunnen ook sprint intervaltraining (SIT) en hoge intensiteit intervaltraining (HIIT) gecategoriseerd worden. De duur van intervaltrainingen varieert tussen de 10 en 90 minuten, waarbij kortere duur vaak samen gaat met hogere intensiteiten. De intervallen in deze trainingen variëren van 15 seconden bij SIT tot 15 minuten bij EIT. spiervezelrekrutering bij deze trainingsvorm hangt bijgevolg dan ook van voorgaande modaliteiten af. Wanneer de intervallen een korte duur hebben maar een zeer hoge intensiteit, zoals bijvoorbeeld HIIT, worden type IIa en type IIx in grote mate aangeworven. Dit uit zich in een snelle glycogeendepletie bij deze SV (Gollnick et al., 1973₂).
3.2.4. Sprinttraining
ST wordt gekenmerkt door zeer korte afstanden variërende van 5 tot 250 meter die aan zeer hoge snelheden worden afgelegd. ST van zeer korte duur (max. 9 sec.) en hoge intensiteit rekruteert voornamelijk type IIx vezels. Bij sprint oefeningen van langere duur (max. 2-3 min.) wordt type IIa hoofdzakelijk aangesproken.
3.3. Aanpassingen aan training
3.3.1. Structureel
Naast de natuurlijke determinanten zoals leeftijd, geslacht en erfelijkheid wordt in een aanzienlijk aantal studies gesuggereerd dat ook training een invloed kan hebben op de spiervezeldistributie (Coggan et al., 1992; Green et al., 1979; harb & Henriksson, 1977; Harber et al., 2012). Training
induceert niet enkel veranderingen inzake spiervezeldistributie. Ook op structureel en morfologisch vlak vinden er heel wat adaptaties plaats op niveau van de SV en spiercel. De manier waarop deze aanpassingen gebeuren is afhankelijk van de intensiteit, volume en frequentie van training. Twee belangrijke uitingen van structurele adaptaties aan training in skeletspieren zijn veranderingen in de volumedensiteit van de mitochondria en capillarisatie.
3.3.1.1.Mitochondriën
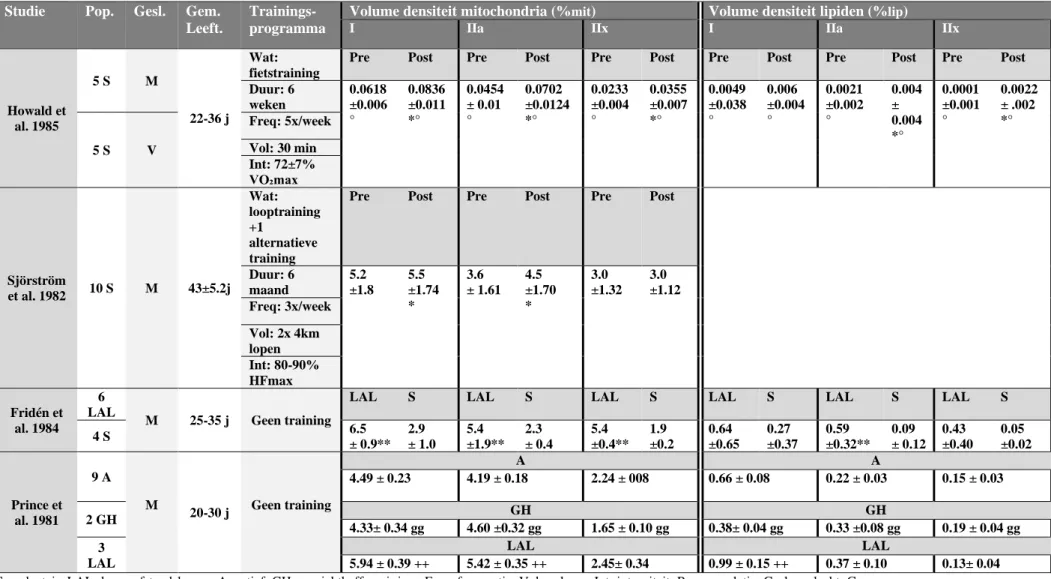
Het mitochondriale volume van skeletspiervezels wordt aangenomen de reflectie te zijn van hun oxidatieve capaciteit. Type I vezels hebben dan ook van nature de grootste mitochondriale inhoud (Howald et al., 1985). DT lijkt een geschikte trainingsvorm om de mitochondriale densiteit in SV te verhogen (zie tabel 3). Een belangrijk gegeven hierbij is dat, afhankelijk van de modaliteiten van DT, deze training een doeltreffende trainingsstimulus zal zijn voor het verhogen van de mitochondriale densiteit in specifieke spiervezeltypes. Waar lange duurtrainingen aan lage intensiteit typisch de mitochondriale inhoud van trage SV zal verhogen, zullen bij hogere intensiteiten ook snelle SV in grotere aantallen gerekruteerd worden. In de studie van Howald et al. (1985) induceert 6 weken DT aan hogere intensiteiten (72 ± 7% VO₂max) een stijging in mitochondriale densiteit van 35%, 55% en 52% in respectievelijk de type I, type IIa en type IIx SV. De kleinere percentuele stijging bij de type I vezels is deels toe te schrijven aan de hogere absolute mitochondriale inhoud bij deze vezels voor training. Daarnaast zorgt de korte duur van deze training voor een niet-optimale stimulus voor deze SV (Howald et al., 1985). Ook HIT is een volwaardige training om veranderingen te induceren in de oxidatieve capaciteit van de skeletspier. Dit is hoogstwaarschijnlijk het gevolg van een hoge mate van spiervezelrekrutering en het potentieel om type II vezels adequaat te stimuleren (Gibala & McGee, 2008).
Bij inspanningen met zeer hoge intensiteiten en korte duur wordt de energie voornamelijk anaeroob geleverd. Hierbij worden preferentieel snelle glycolytische spiervezels gerekruteerd (Prince et al., 1981). Desondanks de hoge rekrutering van type IIx vezels bij gewichtheffen, vertonen de desbetreffende spiervezels in de studie van Prince et al. (1981) een lagere mate van mitochondriale
stijging in de mitochondriale inhoud van type IIx vezels te bewerkstelligen. Een aerobe stimulus lijkt dus met andere woorden onontbeerlijk (Prince et al., 1981; Fridén et al., 1984).
3.3.1.2.Capillarisatie
Opdat mitochondriën optimaal kunnen functioneren, is een adequate aanvoer van zuurstof vereist. Deze zuurstof komt, gebonden aan hemoglobine en myoglobine, via arteriolen en vervolgens via de capillairen tot bij de mitochondriën in de spier. Volgens Laughlin & Roseguini (2008) hebben type I SV van nature een hogere mate van capillarisatie, arteriole densiteit en bijgevolg een hogere oxidatieve capaciteit in vergelijking met snelle SV. Capillarisatie lijkt, net zoals mitochondriale densiteit, toe te nemen in skeletspierregio’s die tijdens training voldoende geactiveerd worden. DT aan lage intensiteit zal bijgevolg voor verhoging in capillarisatie zorgen in regio’s met een hoge concentratie aan type I en type IIa vezels, terwijl er helemaal geen uitbreiding zal zijn van het capillair netwerk in regio’s die voornamelijk uit type IIx vezels bestaan. De logica van het size principle volgend, zal SIT resulteren in een stijging van de capillaire densiteit bij spierregio’s met vooral type IIx vezels (Laughlin en Roseguini, 2008). Duurtraining stimuleert daarnaast ook een vergroting in arteriolaire densiteit in zowel regio’s met veel type I en IIa vezels, als in regio’s met veel type IIx vezels. SIT lijkt geen effect te hebben op de arteriole densiteit. Noch in regio’s met veel type I en IIa, noch in regio’s met veel type IIx vezels (Laughlin et al., 2006). Hieruit kan besloten worden dat enkel DT voor de toename in arteriole densiteit zal zorgen, maar dat zowel DT als SIT toegenomen capillarisatie kunnen teweegbrengen.
3.3.2. Metabool
Metabole adaptaties in de spier worden gereflecteerd door veranderingen in onder andere myosine ATPase (Schluter & Fitts, 1994), enzymactiviteit (Henriksson en Reitman, 1976; Eriksson et al., 1973 ) en substraatverbruik (Howald et al., 1985; Fridén et al., 1984; Prince et al., 1981). Het enzym myosine ATPase hydrolyseert de ATP, hetgeen vastzit aan de myosine kop, in adenosinedifosfaat (ADP) en een fosfaat-ion (Pi). Myosine ATPase speelt bijgevolg een belangrijke rol in de regulatie van de spiercontractie (Goody and Holmes., 1983). Een hoge activiteit van dit enzym lijkt positief gecorreleerd te zijn met een hogere verkortingssnelheid (r= 0.88, p<0.05), wat typerend is voor snelle vezels. Toch lijkt het ook mogelijk om via DT de activiteitsgraad van dit enzym in type I vezels te verhogen. 12 tot 18 weken DT (2uur per dag, 5x/week) zorgde bij de studie van Schluter
en Fitts (1994) voor een toename van 23% in myosine ATPase voor type I SV bij ratten. Bijgevolg werd ook een gelijkmatige stijging in de maximale contractiesnelheid (Vo) van de type I vezels geobserveerd (23%, p<0.05). De submaximale intensiteit van de trainingen zorgden voor een verwaarloosbare rekrutering van type IIa en IIb vezels. De myofibrillaire ATPase activiteit bij deze vezels was derhalve niet veranderd. Canepari et al. (2005) toonden aan dat een stijging in myosine ATPase activiteit in een geïsoleerde SV sterk toenam, zonder dat daarvoor een verandering in de MHC isovorm moet plaatsnemen, wat in meer gedateerde studies werd gesuggereerd (Schluter en Fitts., 1994; Baldwin et al., 1975)
Tabel 3: volume densiteit mitochondria en lipiden
S=sedentair; LAL=lange afstandslopers; A=actief; GH=gewichtheffers; j=jaar; Freq=frequentie; Vol=volume; Int=intensiteit; Pop=populatie; Gesl=geslacht; Gem. leeft=gemiddelde leeftijd
Geen training= cross sectionele vergelijking tussen groepen Studie Pop. Gesl. Gem.
Leeft.
Trainings- programma
Volume densiteit mitochondria (%mit) Volume densiteit lipiden (%lip)
I IIa IIx I IIa IIx
Howald et al. 1985 5 S M 22-36 j Wat: fietstraining
Pre Post Pre Post Pre Post Pre Post Pre Post Pre Post
Duur: 6 weken 0.0618 ±0.006 ° 0.0836 ±0.011 *° 0.0454 ± 0.01 ° 0.0702 ±0.0124 *° 0.0233 ±0.004 ° 0.0355 ±0.007 *° 0.0049 ±0.038 ° 0.006 ±0.004 ° 0.0021 ±0.002 ° 0.004 ± 0.004 *° 0.0001 ±0.001 ° 0.0022 ± .002 *° 5 S V Freq: 5x/week Vol: 30 min Int: 72±7% VO₂max Sjörström et al. 1982 10 S M 43±5.2j Wat: looptraining +1 alternatieve training
Pre Post Pre Post Pre Post
Duur: 6 maand 5.2 ±1.8 5.5 ±1.74 * 3.6 ± 1.61 4.5 ±1.70 * 3.0 ±1.32 3.0 ±1.12 Freq: 3x/week Vol: 2x 4km lopen Int: 80-90% HFmax Fridén et al. 1984 6 LAL M 25-35 j Geen training
LAL S LAL S LAL S LAL S LAL S LAL S
4 S 6.5 ± 0.9** 2.9 ± 1.0 5.4 ±1.9** 2.3 ± 0.4 5.4 ±0.4** 1.9 ±0.2 0.64 ±0.65 0.27 ±0.37 0.59 ±0.32** 0.09 ± 0.12 0.43 ±0.40 0.05 ±0.02 Prince et al. 1981 9 A M 20-30 j Geen training A A 4.49 ± 0.23 4.19 ± 0.18 2.24 ± 008 0.66 ± 0.08 0.22 ± 0.03 0.15 ± 0.03 2 GH GH GH 4.33± 0.34 gg 4.60 ±0.32 gg 1.65 ± 0.10 gg 0.38± 0.04 gg 0.33 ±0.08 gg 0.19 ± 0.04 gg 3 LAL LAL LAL 5.94 ± 0.39 ++ 5.42 ± 0.35 ++ 2.45± 0.34 0.99 ± 0.15 ++ 0.37 ± 0.10 0.13± 0.04
Uit tabel 3 valt af te leiden dat duurtraining voor een toename in intracellulaire lipidenconcentratie zorgt. Dit is niet enkel het geval in de meer oxidatieve SV, maar ook in de glycolytische type IIx vezels indien deze gerekruteerd worden tijdens DT ( Howald et al., 1985). Howald et al. (1985) toonden een toename van 144% aan in de intracellulaire vetinhoud van het laatstgenoemde spiervezeltype na zes weken DT aan hoge intensiteit. Ook de concentratie in de type IIa vezels kende bijna een verdubbeling. Door de aard van de DT in deze studie werd er geen significante verandering bij type I vezels waargenomen. Gevolg van een toegenomen intracellulaire lipidenconcentratie is dat er meer beroep kan worden gedaan op vet als substraat voor de mitochondriale oxidatie tijdens duurinspanningen, met een glycogeensparend effect als primair voordeel (Howald et al., 1985). Wanneer de intracellulaire lipideninhoud van type I vezels bij lange afstandslopers vergeleken wordt met deze van niet specifiek getrainde controlegroepen, kan deze tweemaal grotere proporties aannemen (Prince et al., 1981).
Om de verhoogde intracellulaire lipidenconcentratie te kunnen gebruiken voor mitochondriale oxidatie, is een hoge concentratie van oxidatieve enzymen vereist. Succinaat-dehydrogenase (SDH) is een vaak beschreven voorbeeld van dit type enzymen. Henriksson & Reitman (1976) suggereren een sterk verband tussen spiervezelrekrutering en toename in SDH-activiteit in de verschillende spiervezeltypes. DT aan lage intensiteit resulteerde in een stijging van 32% in de SDH-activiteit van de type I vezels. Adaptatie in SDH concentratie bij type II vezels werd niet gerealiseerd bij deze trainingsvorm. IT gedurende 8 weken (5x vier minuten hoge intensiteit, gevolgd door twee minuten rust) zorgde voor een stijging van 49% in de SDH activiteit van de type II vezels, zonder verandering in de SDH concentratie bij type I vezels. Dat er geen verandering plaatsvond in de type I vezels komt waarschijnlijk door de korte duur van deze trainingen, welke voor een inadequate stimulatie van deze SV zorgt. Hierdoor werden geen adaptaties in oxidatieve capaciteit gerealiseerd (Henriksson & Reitman, 1976). ST aan de andere kant, dat vooral type IIx vezels rekruteert, zorgt voor een toename in de concentratie van het glycolytisch enzym fosfofructokinase bij deze spiervezels. Oxidatieve enzymen zoals SDH nemen in geen enkel spiervezeltype toe in concentratie na ST (Eriksson et al., 1973).
vezels tijdens duurtraining adequaat gestimuleerd worden, deze ook oxidatieve aanpassingen ondervinden na een trainingsperiode (Howald et al., 1985).
3.3.3. Functioneel
De kracht-snelheidsrelatie is cruciaal om een optimaal vermogen te bekomen (Widrick et al.,1996; Cormie et al., 2011). Er bestaat een invers verband tussen deze twee factoren (zie grafiek 2, uit Cormie et al., 2011). Wanneer de snelheid van de concentrische spieractie te hoog wordt, verliest de spier een stuk van zijn capaciteit om kracht te genereren (Widrick et al. 1996). Bij te hoge snelheden komen er minder actine-myosine kruisbruggen tot stand met als gevolg een lagere krachtontwikkeling (Cormie et al., 2011). Om een maximaal vermogen te bekomen, is er dus een evenwicht nodig tussen verkortingssnelheid en krachtproductie.
Grafiek 2: kracht-snelheidsrelatie concentrische spiercontractie skeletspier (uit Cormie et al., 2011)
Zoals eerder gezegd zorgt DT voor het verhogen van de maximale verkortingssnelheid (Vo) bij type I vezels (Schluter en Fitts, 1994). Widrick et al. (1996) observeerden een 19% hogere Vo van de type I bij master lopers (~120 km per week, gedurende 20-25 jaar) in vergelijking met een sedentaire controlegroep. Het absoluut piek vermogen van de type I en type IIa vezels bij de lopers was respectievelijk 13% en 27% lager in vergelijking met hun sedentaire controles. Dit is voornamelijk het gevolg van een daling in dwarsdoorsnede van deze vezels bij de lopers (Trappe et al., 2006; Widrick et al.,1996). Bij continue inspanningen zoals lopen, speelt maximaal vermogen echter slechts een geringe rol.
4. Variatie in respons op training
Heel wat trainingsvormen worden naar voor geschoven als zijnde een geschikte vorm om vooruitgang te realiseren in verschillende cardiorespiratoire parameters. Onder andere duurtrainingen (Howald et al., 1985), HIIT (Astorino et al., 2012), SIT (Willoughby et al., 2016), maar evengoed ook krachttraining (Hickson, Rosenkoetter en Brown., 1980) zouden dit kunnen bewerkstelligen. De grote verscheidenheid aan effectieve trainingsvormen bemoeilijkt de zoektocht naar de meest geschikte training voor een individu om verbetering in de cardiorespiratoire parameters te bewerkstelligen, en bijgevolg prestatie te optimaliseren.
4.1. Probleem van het groepsgemiddelde
Het merendeel van de studies die de metabole, structurele en functionele effecten van een vooropgestelde trainingsvorm met bepaalde trainingsmodaliteiten willen nagaan, doen dit a.d.h.v. een experimenteel design. Vaak is er een interventiegroep waarin de subjecten een bepaald trainingsprogramma volgen en een controlegroep die geen training doet. Een uitspraak over het effect van de training wordt meestal gedaan op basis van het verschil in groepsgemiddelde tussen de twee groepen. In een aanzienlijk deel van deze studies wordt weinig tot geen aandacht besteed aan de interindividuele verschillen in effectgrootte binnen de interventiegroep (Bouchard & Rankinen, 2001). Onder andere de respons in verhoging van het maximale aerobe vermogen, ook wel de VO₂max, lijkt sterk te variëren bij subjecten die eenzelfde trainingsprogramma volgen. Dit valt af te leiden uit Bouchard & Rankinen (2001), waarbij onderzoeksdata uit 3 andere studies werd gebruikt (Bouchard, 1983; Bouchard, 1986; Lortie et al., 1984). Verschillen in VO₂max respons werden daarbij uitgezuiverd voor leeftijd en geslacht. Een gemiddelde stijging na het doorlopen van een trainingsperiode van ongeveer 25% in VO₂max werd bevonden overheen de 3 studies. Een belangrijk gegeven hierbij is echter wel dat sommige subjecten helemaal geen toename konden realiseren, waar anderen bijna een verdubbeling bewerkstelligden. Wanneer er dus uitsluitend naar het groepsgemiddelde wordt gekeken, kan dit een vertekend beeld creëren over de effectiviteit van de training voor ieder individu afzonderlijk.
4.2. Intra-individuele variatie in respons op training bij verschillende
trainingsvormen
Naast de grote interindividuele variabiliteit in respons op training, worden ook grote intra-individuele verschillen in respons gevonden tussen verschillende trainingsvormen. Bonafiglia et al. (2016) toonde aan dat de stijging in VO₂max, submaximale hartslag en wattage op de anaerobe drempel voor een groep recreatief actieve participanten niet significant verschilde op groepsniveau na een SIT trainingsprogramma, wanneer vergeleken met een EDT programma. Wel werd een zeer grote intra-individuele variabiliteit in trainingsrespons op SIT en EDT gevonden. Dit geeft aan dat sommige participanten meer baat hadden bij duurtrainingen aan lagere intensiteiten voor het verhogen van cardiorespiratoire parameters, terwijl andere juist meer progressie maakten in diezelfde parameters door kortere, hoog-intensieve trainingen te doen. Hautala et al. (2006) gingen na of proefpersonen die geen respons vertoonden in VO₂max na twee weken duurtraining, deze VO₂max toch konden verhogen door hypertrofietraining. Subjecten die tot het laagste kwadrant behoorden inzake stijging VO₂max (1 ± 3%, p > 0.05) na duurtraining, realiseerden daarentegen wel de grootste verbetering in VO₂max na een krachttrainingsprogramma (7 ± 5% ,p < 0.001) (grafiek 3). Een noemenswaardig resultaat uit deze studie is dat de subjecten uit het hoogste kwadrant inzake stijging in VO₂max door duurtraining (16 ± 3%, p < 0.001), ook een hoge toename in VO₂max realiseerden na krachttraining. Dit toont aan dat het geen ‘of-of verhaal’ hoeft te zijn.
Grafiek 3: VO₂max respons op krachttrainingsprogramma. Subjecten werden ingedeeld in kwadranten naargelang hun respons op een duurtrainingsprogramma als volgt: 1 ± 3 (Laag), 6 ± 1 (Gemiddeld -), 9 ± 1 (Gemiddeld +), en 16
4.3. Non-responders
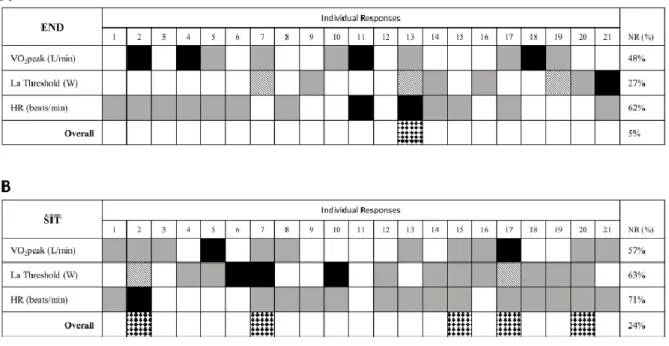
Soms zijn er ook subjecten die helemaal geen respons vertonen op training. Naar deze wordt vaak verwezen zijnde de non-responders. Er bestaat echter geen consensus over het al dan niet bestaan van dit fenomeen. Montero & Lundby (2017) trachtten dit concept in hun studie dan ook te weerleggen. Deze onderzoekers deelden 78 jonge mannen onder in vijf verschillende groepen die respectievelijk 1, 2, 3, 4 en 5 sessies van 60 minuten duurtraining per week uitvoerden, gedurende zes weken. Non-respons na de trainingsperiode werd gedefinieerd als een stijging in maximaal vermogen verkregen uit een maximale inspanningstest (30W/min rampprotocol), kleiner dan de typical error of measurement (TEM). De TEM brengt de meetfout en de dag tot dag variabiliteit in rekening (Hopkins, 2000). In de groepen 1, 2, 3, 4 en 5 waren respectievelijk 69%, 40%, 29%, 0% en 0% zogenaamde non-responders. De subjecten die geen respons vertoonden werden daarna toegewezen aan een identiek trainingsprotocol van zes weken, maar met twee additionele trainingen. Na deze zes weken waren er geen non-responders meer. Deze resultaten suggereren dat wanneer de frequentie van trainingen voldoende hoog is, non-respons kan worden uitgesloten. Ook de resultaten uit de studie van Bonafiglia et al. (2016) zijn in lijn met deze bevindingen. Hierin werden naast de VO₂max ook andere cardiorespiratoire parameters (vermogen op de anaerobe drempel en submaximale hartslag) aangewend om iemand al dan niet te classificeren als non-responder. Indien er alleen gekeken werd naar VO₂max, zouden 10 van de 21 proefpersonen worden geclassificeerd als non-responder na de duurtraining, en na de sprint interval training zouden dit er 12 zijn (zie figuur 2). Men zou op die manier verkeerdelijk kunnen concluderen dat deze proefpersonen niet reageerden op de training. Het is namelijk zo dat wanneer andere cardiorespiratoire parameters geïncludeerd worden, er bij duurtraining slechts één subject voor geen enkele parameter een trainingseffect vertoond. Voor SIT zouden dit er uiteindelijk vier zijn. Een concrete definiëring van non-respons lijkt dus noodzakelijk om verwarring te vermijden.
Figuur 2: Individuele respons na 3 weken duurtraining (A) en 3 weken Sprint Interval Training (B). Positief trainingseffect (witte
blokken), geen trainingseffect (grijze blokken) en negatief trainingseffect (zwarte blokken) worden weergegeven voor de verschillende prestatieparameters. “overall non-responders” worden geïdentificeerd door een met diamant gevulde blok. De kolom NR(%) geeft het aantal non-responders weer voor iedere parameter. (Uit Bonafiglia et al., 2016).
4.4. Oorzaken variatie in respons op training
De rol van genotype op de variatie in cardiovasculaire, metabole en hormonale responsen op aerobe training lijkt van aanzienlijk belang (Bouchard et al., 1999). Om het effect van erfelijkheid te kunnen onderzoeken werden 481 subjecten uit 98 families onderworpen aan duurtraining in de studie van Bouchard et al. (1999). Het primaire resultaat van deze studie is dat er 2.5 keer meer variatie in VO₂max respons op training tussen de families was, dan binnen de families (p < 0.001). Erfelijkheid zou bijgevolg (maximaal) 47% van de variantie, reeds uitgezuiverd voor leeftijd en geslacht, in VO₂max respons op training verklaren (zie tabel 4). Leeftijd, geslacht en initiële Vo₂max leken daarentegen slechts 11% van de variantie van de toename in VO₂max te verklaren (Bouchard & Rankinen, 2001). Erfelijkheid lijkt dus de belangrijkste factor te zijn inzake de individuele adaptaties in VO₂max na een standaard trainingsprogramma. Naast variatie in respons op maximaal niveau (VO₂max), lijkt ook de respons op training op het submaximale niveau vatbaar voor biologische variabiliteit. Na de 20 weken training in de studie van Bouchard et al. (1999) werd een gemiddelde daling van 11 ± 10 hartslagen per minuut waargenomen bij fietsen aan 50W (Bouchard & Rankinen, 2001). Variatie in de daling van de hartfrequentie (HF) bij deze submaximale inspanning lijkt sterk gerelateerd aan de initiële HF. Subjecten wiens initiële HF boven de mediaan lag, hadden na 20
weken een gemiddelde daling van 16 ± 10 slagen per minuut (sl/min), waar subjecten met een initiële HF onder de mediaan slechts een daling van 7 ± 8 sl/min hadden. Initiële HF verklaarde 40 % van de variantie in de daling van de HF na training. Geslacht 5%, ras en leeftijd elk slechts 1%. Het doel van deze scriptie bestaat eruit om na te gaan of de spiervezeldistributie van een subject een mogelijke determinant is voor de variatie in trainingsadaptaties. Er kan worden verwacht dat het individualiseren van trainingsprogramma’s op basis van de spiervezeltypesamenstelling tot een meer homogene respons zal leiden binnen een groep subjecten met een uitgesproken aanwezigheid van een bepaald spiervezeltype.
O=ongetrainden; GR=Geen respons; Wmax= wattage bekomen uit maximale inspanningstest (30W/min); j= jaar; freq= frequentie; vol= volume; M=mannen; V=vrouwen;
Studie Pop. Geslacht
Leeftijd
protocol Initiële VO₂max (gem) VO₂max post Percentuele toename Bereik toename VO₂max Howald et al. 1985 S 5 M 22-36 j
Wat: Fietstraining 43.2 ± 6.3 ml. min−1.kg−1 49.4 ± 5.6 ml. min−1.kg−1 ** 14** /
Duur: 6 weken Freq: 5x/week 5 V Vol: 30 min Intensiteit: 72±7% VO₂max Bouchard et al. 1999 S 95 Vaders 53±5.4 j
Wat: Fietstraining V.: 2618.9 ± 451.1 mlmin−1 V.: 2993 ± 484.2 ml/min V.: 14.3°
***
GR tot 1.01 l/min Duur: 20 weken
86 Moeders 52±4.9 j Freq: 5x/week M.: 1639.5 ± 256.8 mlmin
−1 M.: 1934 ± 287.8 ml/min M.: 18.2°
*** 141 Zonen 25.4±6.1 j Vol: 30 tot 50 min Z.: 3294.2 ± 497.3 mlmin
−1 Z.: 3780.3 ± 517.6 l/min Z.: 14.7° *** 159 Dochters 25.5±6.4 j Intensiteit: 50% tot 75% VO₂max D.: 2063 ± 306.7 mlmin−1 D.: 2433.6 ± 368.4 ml/min D.: 17.9° *** Montero & Lundby. 2017 O 78 M 26.2±3.4 j
Wat: fietstraining Groep 1: 42.4 ± 8.5 ml. min−1.kg−1
1: +0.42 ± 2.4 ml. min−1.kg−1 1: 0.9 /
Duur: 6 weken Groep 2: 44.7 ± 10.7 ml.min−1.kg−1 2: +2.19 ± 2.34 ml. min−1.kg−1* 2: 4.8* Freq: 1,2,3,4 of 5x/week Groep 1: 1x/week Groep 2: 2x/week Groep 3: 3x/week Groep 4: 4x/week Groep 5: 5x/week Groep 3: 43.7 ± 4.2 ml.min−1.kg−1 3: +2.47 ± 1.9 ml. min−1.kg−1 * 3: 5.7*
Volume: 60 min Groep 4: 44.5 ± 5.8 ml.min−1.kg−1
4: +6.5 ± 2.98 ml. min−1.kg−1 *
++
4: 14.6*++ Intensiteit: 65% tot 75%
Wmax uit incrementele inspanningstest (30W/min) Groep 5: 37.4 ± 7.5 ml.min−1.kg−1 5: +10.49 ± 3.9 ml. min−1.kg−1 * ++ 5: 28*++ Hautala et al. 2006 S 73 M & V 42±5 j
Wat: fietstraining 34±7 ml.min−1.kg−1 37±7 ml. min−1.kg−1** 8±6** -5% tot 22%
Duur: 2 weken Freq: 2-3/week Lortie et al. 1984 S 11 M 25±4 j
Wat: fietstraining M: 41± 7 ml.min−1.kg−1 M: 53 ± 7 ml. min−1.kg−1 M: 35** M: 5% tot 46%
Duur: 20 weken
13 V Freq: 4-5x/week V: 33± 4 ml. min−1.kg−1 V: 44 ± 3 ml. min−1.kg−1 V: 41** V: 15% tot 88%
4.5. Bestaande individualisering
4.5.1. Hartslagvariabiliteit
Zoals eerder aangetoond wordt in standaard trainingsprogramma’s enorm veel heterogeniteit inzake de effecten van de training gevonden (Bouchard et al., 2001; Hautala et al., 2006). Hartslagvariabiliteit (HRV) kan als tool gebruikt worden om een DT programma op individueel niveau aan te passen. HRV is de slag tot slag variatie in tijd van opeenvolgende hartslagen. Algemeen genomen kan een daling in HRV niet enkel gerelateerd worden aan verschillende cardiovasculaire condities (Routledge et al., 2010), maar kan het ook in de sportsetting gebruikt worden om overtraining te detecteren (Achten & Jeukendrup. 2003).
4.5.2. Trainingsbelasting
Het kwantificeren van de trainingsbelasting (TL) is essentieel om enerzijds te bepalen of een atleet zich aanpast aan een trainingsprogramma en anderzijds om het risico op ontwikkeling van niet-functionele overbelasting te reduceren (Halson, 2014). Daarnaast geeft het ook de opportuniteit om een beter inzicht te krijgen in de capaciteiten van de atleet om deze dan correct te kunnen bijsturen. TL kan gekwantificeerd worden via interne en externe belasting. Interne belasting steunt op psychologische en fysiologische belasting die een atleet ondervindt tijdens fysieke inspanning. Psychologische bepaling gebeurt via sRPE (session ratings of perceived exertion) aan de hand van een CR-10 schaal (Borg et al., 1987). Dit is een subjectieve score die de atleet toekent aan een trainingssessie en wordt vermenigvuldigt met de duur van deze sessie. Verder wordt er gebruik gemaakt van trainingsimpulsen (TRIMP). Er bestaan verschillende soorten TRIMP’s die afhankelijk kunnen zijn van verandering in HF, lactaatdrempels of een andere intensiteitsfactor (Morton et al., 1990; Lucía et al., 2000; Borg et al., 1987). Externe belasting kan gekwantificeerd worden aan de hand bijvoorbeeld een training stress score (TSS), welke het vermogen gaan meten (Halson, 2014).
5. Trainingsprogramma en spiervezeltype
5.1. Vermoeidheid en herstel
Training omvat een bepaalde belasting die aanleiding geeft tot vermoeidheid. Na deze vermoeidheid treedt herstel op. Dit herstel is interindividueel sterk verschillend en voor een groot deel afhankelijk van spiervezeltypesamenstelling. Na herstel vindt er een door training-geïnduceerde tijdelijk oefeneffect plaats. Hierbij spreekt men van supercompensatie (zie figuur 6). Het toedienen van een trainingsprikkel in deze fase leidt tot het bekomen van een hoger prestatieniveau. Indien er geen nieuwe trainingsprikkel volgt in deze fase verdwijnt het tijdelijk effect en daalt het prestatievermogen opnieuw tot op de beginsituatie. Als de trainingsprikkels elkaar te snel opvolgen en hierbij dus geen volledig herstel plaatsvindt, ontstaat een daling van het functioneel prestatievermogen (Bourgois & Vrijens, 2016).
5.1.1. Vermoeidheid
Vermoeidheid van spieren of SV is een vaak bediscussieerd onderwerp in wetenschappelijke literatuur. Er gebeurden reeds vele onderzoeken naar oorzaken van vermoeidheid bij isometrische (Häkkinnen & Komi, 1986), isokinetische (Komi & Tesch, 1979; Thorstensson & Karlsson, 1976; Komi en Rusko, 1974) en dynamische contracties (Lievens et al., 2020). In deze studies komt er steeds naar voor dat subjecten met een grote proportie snelle SV een hoger piek krachtmoment en/of piek vermogen demonstreren. De dominante aanwezigheid van type II vezels maakt deze subjecten echter meer vatbaar voor vermoeidheid bij herhaalde contracties.
Ook bij vermoeidheid speelt het size principle een belangrijke rol. Häkkinnen & Komi (1986) observeerden tijdens een isometrische vermoeidheidstaak met submaximale weerstand (60% maximale vrijwillige isometrische contractie) een stijging in het geïntegreerd elektromyografisch (iEMG) signaal, wat op een gestegen spieractivatie wijst. Deze stijging in iEMG kan worden toegeschreven aan het falen in de capaciteit om kracht te produceren van de eerst gerekruteerde, kleinere ME tijdens de contractie. Dit falen wordt gecompenseerd door de rekrutering van nieuwe, grotere ME en door de verhoging in de frequentie van de aansturing van actiepotentialen bij de reeds actieve motorneuronen.
Thorstensson & Karlsson (1976) toonden aan dat 50 herhaalde maximale isokinetische knie-extensies, aan een snelheid van 3.14 radialen per seconde, gemiddeld voor een verval van 45 %
tussen de grootte van het verval en het percentage type II vezels bij subjecten (r = 0.86, p < 0.01). Komi & Tesch (1979) zagen verder dat door de aard van dit soort maximale contracties al van bij de eerste contractie alle type IIa en IIx vezels gerekruteerd worden. Bij het herhaaldelijk uitvoeren van maximale isokinetische contracties zal de frequentie van de aansturing van actiepotentialen door grote motorneuronen sneller dalen. Hierdoor kennen subjecten die veel van deze motorneuronen bezitten (snelle groep) een significante daling iEMG signaal (15 ± 3 %, p < 0.01). Bij subjecten met voornamelijk kleine motorneuronen (trage groep) was er na 100 maximale isometrische contracties bij deze studie geen significante daling in iEMG merkbaar (4±3% n.s.). Een stijging in de grootte van de iEMG-signaal/krachtmoment verhouding werd bij beide groepen geobserveerd (p < 0.01). Naast daling in iEMG vertonen subjecten met voornamelijk type II vezels ook een sterkere daling in de snelheid van maximale krachtontwikkeling bij het herhaaldelijk uitvoeren van isometrische contracties (Viitasalo & Komi, 1981).
Uit Lievens et al. (2020) valt af te leiden dat ook bij dynamische contracties subjecten met voornamelijk snelle SV gekenmerkt worden door een grotere vatbaarheid voor vermoeidheid. Subjecten werden ingedeeld in twee groepen o.b.v. hun spiervezeldistributie en voerden drie keer een 30 seconden durende Wingate test uit met telkens vier minuten recuperatie tussen de tests. Het piekvermogen bij de eerste Wingate test was significant hoger in de groep met subjecten die uitgesproken meer type II vezels. Bij de 2e en 3e Wingate test was er geen significant verschil meer waarneembaar tussen de twee groepen. Het totale verval, bepaald als zijnde het verschil tussen het hoogste en laagst gemeten vermogen, over de drie
Wingate tests was groter bij groep die meer snelle vezels had (-61 %) in vergelijking met de type I groep (-41 %) (zie grafiek 4; p < 0.001, uit Lievens et al. 2020).
Grafiek 4: vermoeidheidsprofiel bij ST en FT. ST=groep met voornamelijk trage SV;
FT=groep met voornamelijk snelle SV (uit Lievens et al., 2020)
Ter conclusie kan gesteld worden dat vermoeidheid ontstaat door zowel neuromusculaire, metabole als functionele veranderingen tijdens contractie. Personen met voornamelijk trage spiervezels kennen een minder groot verval in geleverd vermogen of krachtmoment bij maximale inspanningen.
5.1.2. Herstel
Herstel van zowel de snelle als trage SV berust op aerobe processen (Idstrom et al., 1985). Type I vezels lijken onder normale omstandigheden gemakkelijker te herstellen van maximale spiercontracties (Colliander et al., 1988; Lievens et al., 2020). Wanneer er sprake is van bloedtoevoerrestrictie, lijkt het herstel bij de oxidatieve type I vezels hier het meeste onder te lijden. Toch vindt herstel ook onder deze conditie sneller plaats bij deze SV (p < 0.05) (Colliander et al., 1988). In Lievens et al. (2020) wordt het herstel op spierniveau nagegaan door de bekomen kracht uit een maximale vrijwillige isometrische contractie (MVC) in combinatie met elektrostimulatie op vaste tijdstippen na de drie wingates te gaan vergelijken met de bekomen kracht uit de MVC vóór deze testen. Er werden in totaal 8 MVC’s uitgevoerd op verschillende tijdstippen, variërende van tien minuten tot vijf uur na de Wingate testen. Herstel werd bekomen op het tijdstip wanneer de MVC kracht na de Wingate testen niet significant verschilde met MVC kracht van ervoor. Herstel trad bij de groep met voornamelijk snelle SV duidelijk later op in vergelijking met de trage SV-groep. Bij deze laatste groep werd reeds na 20 minuten herstel vastgesteld. De groep met voornamelijk type II vezels was na vijf uur nog steeds niet in staat de initiële MVC kracht te evenaren (Zie grafiek 5, uit Lievens et al.,
iEMG/gemiddeld-krachtmoment verhouding in vergelijking met de initiële MVC. Dit betekent dat ondanks een grotere activatie van de kniestrekkers er niet meer kracht geleverd kon worden. Deze verhouding was niet aangepast bij de trage groep.
Grafiek 5: tijd tot herstel na Wingate testen bij snelle en trage groep (uit Lievens et al., 2020)
6. Onderzoeksvraag en hypothese
Uit voorgaand onderzoek blijkt dat er een grote interindividuele variatie bestaat inzake de VO₂max respons op een bepaald trainingsprogramma. Verschillende parameters zouden hierin een rol kunnen spelen, maar momenteel lijkt enkel erfelijkheid een deel van deze puzzel te verklaren.
Uit wetenschappelijke literatuur blijkt echter dat adaptaties aan training kunnen variëren tussen verschillende SV-types naargelang de duur en intensiteit van de training. DT aan lage intensiteit zal zo bijvoorbeeld vooral voor structurele en metabole adaptaties zorgen bij de trage vezels aangezien deze dominant gerekruteerd worden. Wanneer training echter wordt doorgedreven door zeer lange duur of hoge intensiteit, wordt beschreven dat ook snelle vezels worden aangesproken. Ook lijken er naast de diverse adaptaties aan een bepaalde trainingsstimuli zich ook duidelijke verschillen voor te doen in de ontwikkeling van vermoeidheid en de snelheid van herstel bij de verschillende SV-types.
Het is bijgevolg niet onlogisch om te redeneren dat de spiervezeldistributie van een persoon een belangrijke parameter zou kunnen zijn waarmee rekening dient gehouden te worden bij het opstellen van een trainingsprogramma. Om dit na te gaan zullen in deze scriptie personen met een verschillende SV type samenstelling onderworpen worden aan een zes weken durende HIIT. Op die manier zal in deze scriptie worden nagegaan of er een verschil is in de respons op training tussen individuen met een uitgesproken snelle- en trage SVT voor VO₂max en andere cardiorespiratoire parameters. Hierbij wordt gehypothetiseerd dat de snelle groep
een grotere toename zal kunnen bewerkstelligen, aangezien uit de literatuur blijkt dat snelle SV meer adequaat gestimuleerd worden bij hogere intensiteiten.
II.
METHODE
1. Populatie
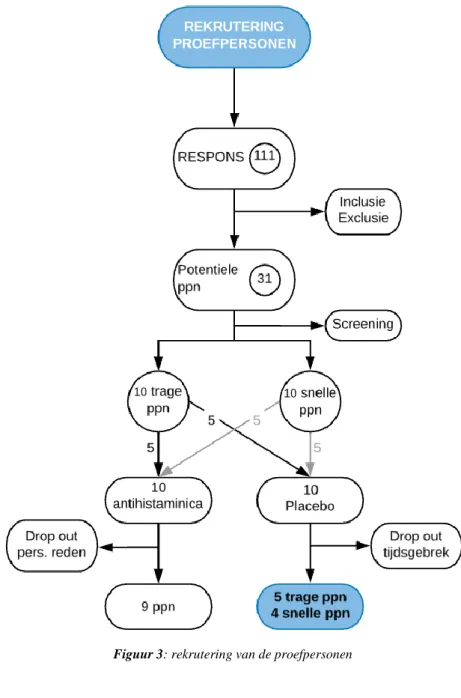
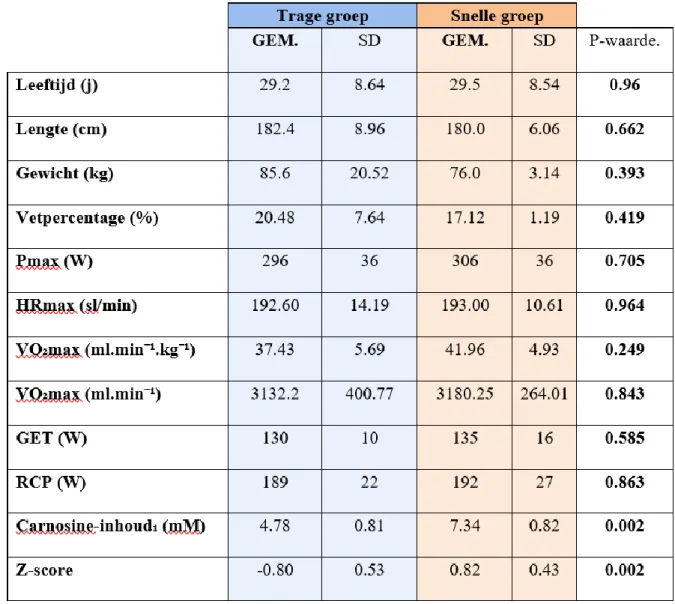
De inclusiecriteria voor deze studie omvatte Kaukasische mannen tussen 18 en 45 jaar die niet fysiek actief waren. Daarentegen behoorden chronische ziekten, het gebruik van medicatie, allergieën, overmatig alcoholgebruik en roken tot de exclusiecriteria. De rekrutering van de proefpersonen kwam tot stand door middel van de verspreiding van een informatieve flyer via sociale media en publieke plaatsen in en rond Gent. Hierop reageerden 111 geïnteresseerden, waarvan 31 nog steeds interesse hadden na verdere toelichting van de studie. Deze werden voorafgaand aan de start van de experimentele studie uitgenodigd voor een eenmalige proton magnetische resonantie spectroscopie (¹H-MRS), hetgeen een inschatting geeft van de spiervezeltypologie bij de geteste subjecten. Vervolgens werden uit de 31 subjecten tien mannen met de meest uitgesproken snelle SV typologie, alsook 10 mannen met de meest trage SV typologie geselecteerd. Deze studie was een combinatie van twee onderzoeksvragen. Voor de onderzoeksvraag die niet aansluit bij dit onderzoek werden de 20 subjecten opgesplitst in twee groepen. Een groep kreeg antihistaminica toegediend, de andere groep een placebo. Door tijdsgebrek en persoonlijke redenen konden twee subjecten, waarvan één uit de antihistaminica groep en één uit de placebo groep, de studie niet voltooien waardoor de uiteindelijke populatie door 18 subjecten werd afgewerkt. Aangezien bij de studie betreffende het antihistaminica supplement geconcludeerd werd dat antihistaminica suppletie effect heeft op de respons op training, werden in de huidige studie enkel subjecten uit de placebogroep geïncludeerd. De uiteindelijke populatie bij dit onderzoek bestond daaropvolgend uit 9 sedentaire Kaukasische mannen (gemiddelde ± standaarddeviatie (SD), leeftijd 29.3 ± 6.7 jaar, lichaamslengte (LL) 181.3 ± 5.8 cm, lichaamsgewicht (LG) 81.4 ± 9.4 kg) waarvan 5 tot de trage groep en 4 tot de snelle groep behoren. Proefpersonen werden gematcht aan andere proefpersonen op basis van LL, LG, vetpercentage en VO₂max door een onafhankelijke onderzoeker. Voorafgaand aan de studie werden alle potentiele risico’s gedetailleerd meegedeeld en een getekend toestemmingsformulier werd verkregen van iedere deelnemer (bijlage 1). Een sportmedisch anamneseformulier (bijlage 2) werd door elk subject ingevuld voor aanvang van de studie en dit formulier werd gecontroleerd door een dokter. Deze studie werd goedgekeurd door het Ethisch Comité van het UZ Gent.
Figuur 3: rekrutering van de proefpersonen
2. Experimentele procedure
2.1. Protocol
¹H-MRS
Voor aanvang van de studie onderging ieder van de 31 subjecten een ¹H-MRS scan. Aan de hand van de nucleaire magnetische resonantie scanner (3 Tesla whole body MRI, Siemens Trio, Erlangen) wordt op basis van magnetische resonantie spectroscopie (MRS) een spectrum